
|
Tiger Predation
|
|
Page 168 to 171 of "Hulirayana Akashavani".
June 1991 - T-03 aka "Dhasa" in action On the morning of June 20, 1991, I went on the trail of a male tiger named Dhasa (T-03). Madhusudan was a college student with me. Madhu has a keen interest in wildlife. That's why I encouraged him to study biology. He is very interested in this field. He would come to the woods with me whenever possible. The same lad today, Dr. M. D. Madhusudan, is a renowned wildlife scientist. By 8am this morning, Dhasa had decided to stop his journey and relax. Recognizing the radio signals coming from Dhasa's collar from three directions, we identified the spot where he was sleeping. Dhasa had chosen the space between the ocean floor and the adjacent fireline for his rest. We walked carefully but told Madhu that we could go as far as fifty meters to the sleeping tiger. The most thrilling experience is to follow a tiger on foot in a dense forest. He was just as excited as I was. The forest department fires along the edge of the forest to prevent forest fires. Have listen carefully, Dhasa radio signals are on this tracks. Something along the edge of this path should be covered with bamboo and mature grass. The sun's rays were spreading out from inside the forest. The clichs of birds that filled the forest around my client was unheard of. The only beeping sounds of the radio collar were reaching my windows. Dealing with all the noise around this deaf forest is just as inviting as danger. In this forest, where there are a couple of elephants per square kilometer, I have only been deafened by an ear deafening warn from an elephant. In a few minutes we came to a small huddle that had been cleared. Radio signals signal the tiger to be about sixty meters away from us, within the bamboo lining of the edge. The sight of the tiger was a different sight. Between the edge of the plain and a tiger bamboo stick was a female Gaur (They named her Katy). Katy glanced at the tiger's hiding spot and never noticed us coming 50 meters away. Katy, who was uniformly eyeing the tiger's hideout, often seemed to be aggressive, hitting her leg and slapping her tail. It looked like she was worried. Katie's attitude made us feel extraordinary. If the tiger is so close, why didn't Katy run away? Why is it that she has left her group's defense and is alone? Looking at it from my binoculars, I knew what was true. A few seconds before we got there, the tiger had attacked. Then all the other members of the group must have fled. Katy's back was severely damaged by the tiger's claws. Blood flowed from her head like a stream. Dhasa also bit Katy's muzzle. Despite being so badly injured, the mighty tiger had to retreat from her. Without killing Katy immediately, Dhasa retreated into the bushes. Dhasa is now looking at his victim. If the gaur tries to run from there, the tiger will attack from behind and bring her to the ground. Thus, at the last moment of her life, Katy showed great courage in spite of her helplessness. Despite the injuries, Katy was a strong rival to a tiger-like enemy. Even if the tiger fails, the slightest mistake is enough, and the strong edges of the strong body, which weighs 600 kilos, are capable of ripping the tiger's stomach. Thus the hunter and the victim got into conflict. For about five minutes we could have seen this thrilling forest drama where a hunter and its victim were in conflict. Curious as to what's ahead of us, I was surprised to find that the radio signals that were coming from the tiger's side suddenly quit the activity and became inactive. Hearing this signal means that the tiger is asleep completely. Can the tiger muscles keep up as it prepares for its final attack on Katy? Will Dhasa sleep and rest from the exhaustion of the first round of fighting such a strong rival? Dhasa had a tendency to catch gaurs (he was a gaur expert). Two months ago, near Kallu Korai Road, Dhasa, had killed a small male Katti, the size of a female. The tiger had no hurry to catch its prey. Dhasa was waiting for the time. There was no group protection for the injured Katy; It doesn't even exist. Katy's condition rapidly deteriorated due to bleeding. The cow's head, which slowly collapsed to the ground without stopping at the end, was visible only to us from the grass. We both got a little closer to looking at Katy better. It was my belief that the collapsed Katy could not stand up but left us in a hurry! The tiger's radio signals, which had been dormant for so long, were suddenly dynamic. It is not known if the tiger moved back and stood up to notice the plummeting Katy. No wonder, though, that fatigue went off in sleep. But this move of the tiger shocked Katy. Immediately Katy stood up and started to cry, tearing her nostrils. Katy had found us who had come close to thirty meters in this chaos! Attirukku turned to us without hesitation to think. Not expecting the most unexpected invasion of Katy, I rode back on the grass. "Run, run," shouted Madhu. There was no need to say so. The long-legged Madhu crossed over to me and pushed forward. We somehow escaped into the woods with antenna cables, receivers and binoculars all around us. We ran back a hundred meters and stood up and looked back. Katy didn't come after us. The dull-eyed Katy could not follow us in the green of the forest. But Katie knew that there was a danger behind her. It is impossible to say what Dhasa might have felt when he saw this strange scene. Can you imagine a funny scene in which two men busting around antennas and cables are driving like madman in a mad rush? Katy's corpse was spotted the next morning when we went to the place. Katie had walked about three kilometers from where she had driven us the previous day. Katy's dull eyes look nothing like dark. The tiger Dhasa, who could see well in the dark, easily caught his prey. The tiger, who seemed to have all the perseverance, eventually ate only about 20 kg of meat and left. There was no hint of Dhasa radio signals anywhere. An hour later, Dhasa's footprints were spotted on the soft soil of Chikkapala Road, one and a half kilometers away. The introduction of Dhasa's footprints was good for me. His footprints are small compared to his body size. But there were also dried blood stains near the footprints. That is, Katy had slightly injured the tiger in the fight the previous day. But the wound was not so great. Dhasa succeeded in killing Katy, ate too much and walked away comfortably. (Dhasa's pug mark) (Katy's carcass being measured)
@Pckts, So, Ladli’s 3 male cubs have dispersed. Good to see. In other Ranthambore news, Arrowhead littered again with 3 cubs (father is most likely T-86 again). Whereas, Forest Department plans to relocate Riddhi (T124) to Sariska as the two sisters and mother have had an endless battle for territory. Riddhi in fact had to be treated with 11 stitches on her tongue after a fight with Siddhi. She wouldn’t have made it without the intervention.
@Khan85, Dhasa or Das (named after the camp cook who was a potly man) was the gaur specialist that Karanth always mentions on the topic of tiger vs Gaur. He was a chunky male tiger who took down quite a few of them. He weighed 258 kilos (if I remember correctly). A bull gaur did kill him eventually. (07-27-2021, 12:25 AM)Ashutosh Wrote: @Pckts, So, Ladli’s 3 male cubs have dispersed. Good to see. In other Ranthambore news, Arrowhead littered again with 3 cubs (father is most likely T-86 again). Whereas, Forest Department plans to relocate Riddhi (T124) to Sariska as the two sisters and mother have had an endless battle for territory. Riddhi in fact had to be treated with 11 stitches on her tongue after a fight with Siddhi. She wouldn’t have made it without the intervention. It was said that T8 was with the Cub, possibly helped with the kill?
Tiger with Gaur kill - By Kartik Mahajan
 *This image is copyright of its original author
Kolsa female with gaur kill:
 *This image is copyright of its original author https://twitter.com/malllige/status/1277390272867913728
12 year old male tiger 'Ravan' with Gaur kill in Satpura:
 *This image is copyright of its original author  *This image is copyright of its original author  *This image is copyright of its original author  *This image is copyright of its original author  *This image is copyright of its original author https://www.facebook.com/156539381083943...610398242/
Diet of Bengal Tiger in Chitwan National Park
ABSTRACT We studied the diet of the Bengal tigers (Panthera tigris tigris) in Chitwan National Park, Nepal, by identifying 109 prey items from 85 tiger scats. Tigers in this region fed upon eight different mammal species. Chital (Axis Axis)was the major prey with a frequency of 45% of the Tigers’ diet. The occurrence of other prey species included Sambar (Cervus Unicolor, 23%), Wild Pig (Sus Scrofa, 15%), Hog Deer (Axis Porcinus, 9%), Barking Deer (Muntiacus Muntjak, 4%) and Gaur (Bos Gaurus, 2%). Tigers also hunted livestock, but this prey comprised a small component of the relative biomass (buffalo 5% and cow 2%). Our study suggests that the tiger depends mostly upon wild prey for its subsistence in the Chitwan National Park, but will also sporadically hunt livestock.  *This image is copyright of its original author
Prey Selection and Food Habits of Large Carnivores: Tiger Panthera Tigris, Leopard Panthera pardus and Dhole cuon alpinus in Mudumalai Tiger Reserve, Tamil Nadu
Prey utilization by tiger as shown by scat and kill data in Mudumalai Tiger Reserve  *This image is copyright of its original author
Prey preferences of the tiger Panthera tigris Abstract Tigers Panthera tigris continue to decline despite the best efforts of the worldwide scientific and conservation communities. Prey depletion has been linked to this decline, but a clear definition of what constitutes preferred prey and preferred prey weight range does not exist. This is critical information if we are to assess tiger reintroduction potential, monitor unforeseen poaching of predators and prey, and successfully conserve the species. Here we reviewed the available literature on tiger diet and prey availability and calculated Jacobs’s electivity index scores from 3187 kills or scats of 32 prey species. We found that wild boar and sambar deer are significantly preferred by tigers, with red deer and barasingha likely to be significantly preferred also with a larger sample size. Prey body mass was the only variable that related to tiger prey preference with species weighing between 60 and 250 kg preferred by tigers yielding a ratio of predator to preferred prey of 1:1, which is similar to other solitary felids. This information can be used to predict tiger diet, carrying capacity and movement patterns, as it has been for Africa’s large predator guild, and has important implications for tiger conservation throughout its distribution. Introduction Tigers Panthera tigris are the largest of the felids with a distribution that once spanned Asia and much of the diverse array of habitats therein (Nowell & Jackson, 1996). Today, tiger numbers have plummeted and its range has vastly contracted (Fig. 1) such that the species is listed as endangered (IUCN, 2009) despite long-term conservation actions, such as the 40-year-old Project Tiger initiative in India (Panwar, 1987) and intensive research. The causes of this decline have been illegal hunting, often for tiger bone, a declining prey base, and habitat loss and degradation (Karanth & Stith, 1999; Carroll & Miquelle, 2006; Chapron et al., 2008; Wilson & Mittermeier, 2009; Miquelle et al., 2010). In light of the importance that prey has for tiger conservation (Karanth & Stith, 1999; Miquelle et al., 2010), it is critical to understand tiger predation to ensure an adequate prey base. There are numerous excellent studies that have documented the local diet of tigers (see examples in Tables 1 and 2) and their local preferences (Støen & Wegge, 1996; Miquelle et al., 2010); however, these are invariably affected by site-specific factors such as variable prey abundance and habitat features that do not afford conclusions to be drawn on the species as a whole compared with a broader meta-analysis. These studies provide data to holistically review the diet and prey selection of the tiger – information that can ultimately be applied to predicting diet, carrying capacity and home range at unstudied sites, as has been done with Africa’s large carnivore guild (Hayward, O’Brien & Kerley, 2007b; Hayward et al., 2007a, 2009). Tigers are solitary, ambush hunters that also actively search for prey (Wilson & Mittermeier, 2009) and require >5 kg of meat daily to maintain body condition (Sunquist, 1981). They are capable of capturing and killing large prey, including adult male gaur Bos gaur (Karanth & Sunquist, 1995), but also take prey as small as hares (Johnsingh, 1983). Tiger diet is biogeographically diverse (Miquelle et al., 1996, 1999; Sunquist, Karanth & Sunquist, 1999). Based on the breadth of their diet across their range, one might infer that tigers are non-selective predators; however, it seems likely that the species’ morphology and solitary hunting strategy imposes limitations on the prey it can capture most efficiently with minimal risk. Solitary leopards P. pardus, cheetahs Acinonyx jubatus and Eurasian lynx Lynx lynx preferentially prey on species at or just below their own body mass (Jedrzejewski et al., 1993; Hayward et al., 2006a,b), whereas group hunters, like lions P. leo and African wild dogs Lycaon pictus preferentially prey on species much larger than themselves (Hayward & Kerley, 2005; Hayward et al., 2006c). Sunquist (2010) has suggested that the size of prey preferred by tigers is approximately their own size, as has been found with other solitary carnivores. Consequently, we aimed to determine the preferred prey of tigers from throughout the species’ range and then test whether these preferred prey species conformed to the prediction that solitary hunters preferentially prey on species of similar body mass to themselves. Methods We determined tiger prey preferences following the methods used to determine the prey preferences of Africa’s large predator guild (Hayward et al., 2006b,c). Specifically, we reviewed the literature on tiger ecology and extracted data on tiger diet and the abundance (actual or relative) of prey at each site. This literature survey revealed 24 published studies from 18 different conservation areas in 7 countries describing the diet of the tiger, which included some measure of prey abundance [either actual or relative (Table 1; Fig. 1)]. Several of these studies were conducted over a long term and these allowed temporally separated prey preferences to be calculated as prey abundance changed over time (Table 1). Such partitioning has been used previously in the study of carnivore ecology (see Creel & Creel, 2002). Therefore, a total of 28 assessments of prey preference were calculated from sites throughout the extant range of the tiger. We do not consider autocorrelation to be problematic by using data from the same area at different levels of prey abundance, as one of the fundamental rules of whether an individual is captured and killed is the probability of the predator encountering it, and this varies with prey density (Hayward & Kerley, 2005). The dietary data collected in these studies were largely derived from incidental observations of kills, although faecal analysis was also used (Table 1). Continuous observations are widely regarded as the superior method of ascertaining the diet of a large predator (Bertram, 1979; Mills, 1992); however, these are extremely difficult with such secretive and elusive predators as the tiger. Global positioning system telemetry locations are probably the best source for determining the diet of solitary carnivores (e.g. Swanepoel, Dalerum & Van Hoven, 2010; Tambling et al., 2010). While other methods may exhibit biases, we are not sure to what extent this may affect these results; however, the tested predictive accuracy of these methods suggest their effect is minor (Hayward et al., 2007a,b). Faecal analysis is another valuable method used in ascertaining predator diets, although if used alone may overemphasize the importance of small prey items (Mills, 1992). Table 1 Sites and sources of prey preference data used in this study  *This image is copyright of its original author  *This image is copyright of its original author Table 2 Preference status, mean Jacobs’s index value of each tiger prey species, number of studies recording it as potential prey (np) and actual prey (na), mean percentage abundance and kills of each prey species, body mass (three-fourths adult female), and categories of herd size and main habitat based on Nowak (1999) and references in Hayward et al. (2006a)  *This image is copyright of its original author  *This image is copyright of its original author The inclusion of studies using all these methods ensured that the majority of prey species of the tiger were assessed in our analysis. Numerous studies provided excellent descriptive information on tiger diet but insufficient or no information on prey abundance (Sankar & Johnsingh, 2002; Talukdar, 2002; Avinandan, 2003), whereas others had no kill data (Reza, Feeroz & Islam, 2002; Shrestha, 2004; Adhikari & Khadka, 2009; Siddiqui, 2010) or were collected over such a long period that great fluctuations in prey abundance occurred (Stephens et al., 2006; Miquelle et al., 2010). This kind of analysis is reliant upon the quality of the underlying data and this may vary from source to source; however, the predictive ability of the methods we employ here and the fact that the studies we cite have been peer-reviewed, suggests the impact of this variation is minor. We used Jacobs’s index to determine the prey selectivity of tigers via the formula: D r p r p rp = ( ) − ( ) + − i i i i ii , 2 where, ri is the proportion of species i among the total kills at a site and pi is the proportion of species i in the available prey community (Jacobs, 1974). The resulting values range from +1 (maximum preference) to -1 (maximum avoidance) (Jacobs, 1974). The mean Jacobs’s index value for each prey species across studies was calculated [ 1 standard error (se) wherever the mean is shown], and these values were tested for significant preference or avoidance using t-tests against an expected value of 0 if they conformed to the assumptions of normality. Where transformation could not satisfy these assumptions, the sign test (Zar, (1999)) was used. There are three major benefits of this kind of meta-analysis. Firstly, it is not biased by the results from one particular area. Secondly, it is not influenced by the available prey community because for a species to be significantly preferred or avoided, it must be so at several sites. Lastly, it is not biased by predation of particular sexes or age classes as the data arise from hunting by tiger populations that consist of both sexes and all age classes. The number of species with relatively small sample sizes (i.e. few studies recording them as prey) means that significant preference and avoidance is less likely because at least five Jacobs’s index values are required to obtain a significant result using the sign test, hence it is a conservative measure of prey preference. Nonetheless, plots of Jacobs’s index with error bars illustrate which species are likely to be significantly preferred or avoided with a larger sample size, assuming the existing trend continues. Multiple regression was conducted on non-correlated, transformed variables to determine which factors influenced the prey preferences of the tiger. The variables used were prey relative abundance at a site, prey body mass, herd size and preferred habitat type (Table 2). Significant relationships were plotted using distance-weighted least-squares and linear regression. Linear regression was also used for testing the relationship between prey relative abundance with Jacobs’s index value and the proportion of kills at a site. We used three-quarters of the mean adult female body mass of prey species to take account of calves and sub-adults eaten. This value was used in previous studies (Hayward et al., 2006b,c), and we continue its use here to allow comparison between these studies. Prey weights were taken from Nowak (1999). We used herd size as an indicator of the ability of prey to detect predators and vice versa. This was a categorical variable, with 1 relating to solitary individuals, 2 to species that exist in pairs, 3 to small family grouping species, 4 to small herds (10–19) and 5 to large herds (20; Table 2). Obviously, this is a simplification as large herding species may also have solitary males among them and species normally existing in smaller groups may aggregate in certain habitats or following habitat disturbance (Schaller, 1967; Dhungel & O’Gara, 1991; Olson et al., 2005); however, this variable has been successfully used previously (Funston, Mills & Biggs, 2001; Hayward et al., 2006c). Habitat type may influence predation rates as the density of vegetation can affect the detectability of both predator and prey. Animals inhabiting dense vegetation generally adopt a silent, solitary, hider strategy to evade detection, whereas prey on open grasslands are detected by sight rather than sound and often exist in large herds (Geist, 1974; Leuthold, 1977). Furthermore, tigers are solitary ambush hunters reliant upon cover to stalk their prey (Wilson & Mittermeier, 2009). On this basis, we would expect solitary tigers to predominately hunt prey inhabiting denser habitat types. Although inherently difficult to classify (Sunquist & Sunquist, 1997), a categorical variable of habitat was used with 1 relating to species living in open environments, 2 to those in woodlands and 3 to those inhabiting dense forests (Table 2). Means are presented with standard errors throughout. Results We calculated Jacobs’s index scores based on 3187 kills or scats of 32 species recorded as tiger prey in the literature (Table 2). These studies had a mean sample size of 120 25 scats or kills (Table 1). Excluding species with just one Jacobs’s index estimate, the mean number of preference estimates per prey species was 8.0 ( 1.6). The most frequently preyed upon species by tigers were wild boar, chital and sambar and these were killed wherever they were available (Table 2). Although less frequently present at study sites, red deer, roe deer and muntjac were also almost invariably killed by tigers (Table 2). Conversely, peafowl, musk deer, goral and Indian rhinoceros were generally avoided when they were present in a community (Table 2). Sambar and chital deer constituted the highest proportion of tiger prey in all sites where they occurred, both comprising 34 (4%) of kills where they occur (Table 2). Red deer, banteng and wild boar were the next most important prey in those locations where they occurred (Table 2). Chital and red deer were the most abundant prey where they occurred (Table 2). Tigers significantly prefer to prey upon wild boar (t54 = 4.38, P < 0.001) and sambar deer [t42 = 4.25, P < 0.001; (Table 2; Fig. 2)]. Red deer and, possibly also, barasingha are likely to be significantly preferred prey of tigers with a larger sample size (Table 2; Fig. 2). Tigers significantly avoided preying on peafowl (Z = 100, n = 6, P = 0.001), langur (t26 = -15.06, P < 0.001) and nilgai (t32 = -2.35, P = 0.025; Table 2; Fig. 2). Larger sample sizes of Indian rhinoceros, common goral, tahr, musk deer, livestock, macaque monkeys, elephants and chinkara gazelle are likely to see them significantly avoided also (Fig. 2). The remaining prey species are killed by tigers in accordance with their availability in the prey community (Fig. 2). A multiple linear regression on prey relative abundance, body mass (log transformed), herd size and main habitat (Table 2), was not significant (R2 = 0.34, F4, 15 = 1.89, P = 0.16). Prey body mass (log) was the only variable that was significantly related to the Jacobs’s index value (t15 = 2.32, P = 0.03). There were positive significant relationships or trends between the proportional abundance of common tiger prey species and the proportion they constituted among kills at a given study site (Fig. 3). There was no relationship between proportional abundance and the Jacobs’s index value of these species; however, there was a negative trend for the two significantly preferred prey species: sambar and wild boar (P < 0.10 for both; Fig. 3). Figure 2 Mean Jacobs’s index values ( 1 standard error) for species that were potential prey of tigers at two or more sites. Black fill illustrates significantly preferred prey species, dark grey species that are likely to be significantly preferred with larger samples sizes, pale grey species as killed in accordance with their proportional abundance by tigers and open bars represent significantly avoided species.  *This image is copyright of its original author When all prey species with >2 estimates of Jacobs’s index were considered, tigers prefer prey weighing between 60 and 250 kg (Fig. 4); however, tigers increasingly prefer prey up to 200 kg (r2 = 0.77, n = 17, P = 0.001; Fig. 5). The mean body mass of preferred prey species, that is those species with two or more Jacobs’s index estimates where the mean (1 se) exceeded 0, was 140 13 kg, and the mean body mass of significantly preferred (ideal) prey species was 130 21 kg (Table 2). On the basis that an adult tigress weighs on average 125 kg (lower range of mean adult female body mass reported for Bengal tigers by Wilson & Mittermeier, 2009), the ratio of predator body mass to that of their preferred prey was 1:1.1 and that of their ideal prey was 1:1. Discussion This study provides information on the key requirements necessary for a source population of tigers to persist; namely, a viable population of large deer (e.g. sambar, barasingha or red deer) and/or wild boar. Tigers prefer preying upon species similar in body mass to themselves akin to the preferences of other solitary predators (Gittleman, 1985; Vezina, 1985; Jedrzejewski et al., 1993; Carbone et al., 1999; Hayward et al., 2006a,b). These preferences are reinforced throughout the predatory behavioural sequence (Creel & Creel, 2002; Hayward et al., 2011) and have theoretically evolved, through optimal foraging (Goss-Custard, 1977; Bertram, 1978; Krebs, 1980), to maximize energetic returns while minimizing injury risk (Hayward & Kerley, 2005). The preferred prey species supports the theory that tiger evolution followed the Pleistocene radiation of cervids in south-eastern Asia as the evolution of large ungulates created a niche for a large-bodied forest-edge predator (Sunquist et al., 1999) and largely quantifies Sunquist’s (2010) preferred prey weight range of tigers. This preferred prey weight range also includes the young of larger species, which are frequently preyed upon, such as rhinoceros calves (Talukdar, 2002) as well as Pere David’s deer Elaphurus davidianus whose behaviour suggests it may have been an important prey item of tigers before its 1200 years of captivity (Li et al., 2011). The preferred prey weight range also accords with that found at well-studied individual sites, like Nagarhole (Karanth & Sunquist, 1992, 1995). It is becoming increasingly evident that preferential predation on suids is practised solely by the largest felids, possibly because of their possession of retractile claws (this study; Hayward & Kerley, 2005; Miquelle et al., 2010). Suids, including wild boar and warthogs Phacochoerus africanus, possess defensive weapons that can inflict serious injuries to predators (Miquelle et al., 2010). This means that predators must avoid these weapons in the act of killing them. Felids are able to effectively do this by using their claws and forepaws to manipulate the suid to keep the tusks for goring them while obtaining a safe killing bite. Suids also possess a low centre of gravity that reduces the effectiveness of coursing predators 2009).who tend to rely on tripping fleeing prey and killing them before they can defend themselves. The tiger’s preference for wild boar is likely to be increased with the inclusion of data from Sumatra, which lacked abundance data, but where wild boar composed a large proportion of the diet (Table 1). The importance of suids to tigers has not previously been identified outside the Russian Far East, but has important implications for conservation actions in all tiger range states. Megaherbivores appear to avoid top-down limitation through preferential predation by large predators, as they do in Africa (Sinclair, Mduma & Brashares, 2003; Hayward, Tigers also provide more evidence (Fig. 3) that prey species that are rare in the environment avoid top-down limitation from predators through their scarcity in the prey community (Hayward, 2011). Karanth & Sunquist (1995) concluded that selective predation in Nagarhole facilitated coexistence of large predators. Leopards in Asia preferentially prey on chital Axis axis and prefer prey weighing between 10 and 40 kg (Hayward et al., 2006a), which is much smaller than prey preferred by tigers (this study). Thus, there is clear separation between leopard and tiger dietary preferences enabling their coexistence. Prey availability and body mass were key determinants of prey preferences of tigers in Nepal’s Royal Bardia National Park (Støen & Wegge, 1996). Wild boar live solitarily in the dry season and were more vulnerable to a stalking tiger (ambush predator) than the large herds of chital (Støen & Wegge, 1996). Johnsingh (1992) supported the theory that the grouping behaviour of chital reduced predation from tigers; however, the results of this present study question this explanation and suggest that chital are too small to be optimal prey of tigers. Tiger density has previously been predicted based on the biomass or density of all potential prey species present at a site, although the confidence intervals are wide (Miquelle et al., 1999; Sunquist et al., 1999; Karanth et al., 2004). Similar relationships exist for large African carnivores and all large carnivores (van Orsdol, Hanby & Bygott, 1985; Laurenson, 1995; Stander et al., 1997; Carbone & Gittleman, 2002). These relationships have been refined for African carnivores by relating predator density with the biomass of their preferred prey rather than all available prey, which has greatly reduced the amount of variation in the data (Hayward et al., 2007b). Similar attempts have been recently made for tigers; however, these are based on a limited number of local studies (Miquelle et al., 2010). It is highly likely that using the results of our study would further improve the accuracy of tiger density estimates over a broader area and more accurate predictions of tiger density could allow the identification of undetected poaching keeping tigers below carrying capacity or be used to improve the accuracy of population viability analyses (Hayward et al., 2007b; Bach et al., 2010). The use of preferred prey biomass could also be used to predict tiger diet and home range size, as has occurred with lions, leopards and spotted hyaenas Crocuta crocuta (Hayward et al., 2007a, 2009).
Indo-chinese tiger Predation Studies
Study of the Indochinese tiger (Panthera tigris corbetti Mazak, 1968) population and habitat use in Phu Khieo Wildlife Sanctuary, Chaiyaphum province [2001] "Prey species found in tiger dung, included : Common Barking Deer (Muntiacus muntjak (Zimmermann) 1780), Common Wild Boar (Sus scrofa Linnaeus, 1758), Sambar Deer (Cervus unicolor (Kerr) 1792), Laesser Mouse Deer (Tragulus javanicus (Osbeck) 1765) and rats (Muridae). Common Barking Deer constituted over 40 percent of prey." Ecological Covariates at Kill Sites Influence Tiger (Panthera tigris) Hunting Success in Huai Kha Khaeng Wildlife Sanctuary, Thailand The Number of Tiger’s Kills and Biomass of Species in 150 Kills in Huai Kha Khaeng Wildlife Sanctuary, Thailand.  *This image is copyright of its original author Weights of gaur ( Bos gaurus ) and banteng ( Bos javanicus ) killed by tigers in Thailand FIGURE 1 Two of the largest prey of tiger in Huai Kha Khaeng (a) Male tiger feeding on gaur (Bos gaurus). (b) Female tiger and her cubs feeding on banteng (B. javanicus)  *This image is copyright of its original author TABLE 1 Mean weights of gaur (Bos gaurus) and banteng (B. javanicus) killed by tigers based on number of kills in different sex and age classes and published weights of those classes from Ahrestani (2018) and Hoogerwerf (1970)  *This image is copyright of its original author TABLE 2 Percentage and numbers of sexes and age classes of gaur (B. gaurus and B. Javanicus) killed by male and female tigers  *This image is copyright of its original author Sumatran Tiger predation Predicting preferred prey of Sumatran tigers (Panthera tigris sumatrae) via spatiotemporal overlap "We documented , captures of potential prey species, with red muntjacs being recorded most frequently, followed by pig-tailed macaques and mouse deer (Table ). Tigers exhibited a diurnal activity pattern (Fig. ) and had the high- est temporal overlap with wild boar, followed by pig-tailed macaques and sambar (Fig. ,Table ). The highest spatial overlap was also with wild boar, followed by sambar and pig-tailed macaques (Table ). When plotting the values indicating spatial and temporal overlap of tigers with potential prey species, wild boar and sambar deer fell in the upper right quadrant, suggesting they are potentially preferred prey. Tapirs were in the lower left quadrant, indicating they were probably not preferred, and the other prey species were in the upper left (high temporal but low spatial overlap), indicating potential alternative prey (Fig. ). Tigers had the greatest spatio-temporal overlap with wild boar, with a spatial and temporal composite score of ., which is % higher than the species with the second highest scores (sambar and pig-tailed macaques, both . Assessing tiger–prey interactions in Sumatran rainforests Large carnivore habitat selection should focus on the most profitable patches, where the lowest levels of energy are expended on searching for and then capturing the largest bodied prey with the least risk (Stephens & Krebs, 1987; Scognamillo et al., 2003; Carbone, Teacher & Rowcliffe, 2007). Thus, the Sumatran tiger would be predicted to select sambar (185–260 kg) and tapir (250–540 kg, Boonsong & McNeely, 1988). For sambar, which appeared to be less common in the KS study areas, there were insufficient data to determine activity patterns confidently. However, similarly designed camera-trap studies from southern Sumatra and Peninsular Malaysia found sambar to have predominantly crepuscular activity patterns (T. O'Brien, unpubl. data; Laidlaw & Shaharuddin, 1998). The high overlap between tiger and muntjac in our study was strong because both species exhibited peaks of activity around dawn and dusk. To date, evidence of interactions between tiger and tapir is limited to photographic records of tiger attacks on tapir and speculation over the tapir's status as a prey species (Lynam, 1999; Holden, Yanuar & Martyr, 2003). Even though the tapir was frequently photographed and along trails used by tiger, our analysis found only a low level of temporal overlap; tapir was predominantly nocturnal. Thus, the lack of a tiger–tapir interaction may be because tapir is not a principal prey species, and this lack of relationship is suggested from Malaysia where overlap was low (Kawanishi & Sunquist, 2004). This is surprising because the Bengal tiger, which although larger than the Sumatran tiger (adult males of 180–258 and 100–140 kg, respectively, Nowell & Jackson, 1996), typically kills not only large prey (>176 kg) especially adult sambar but also occasionally adult male gaur Bos gaurus, which can attain an upper body mass of 1000 kg (Karanth & Sunquist, 1995; Andheria, Karanth & Kumar, 2007). Malayan tapir should not, therefore, be too large for a Sumatran tiger to kill. An alternative explanation for the lack of positive tiger–tapir interaction may be the effect of predation risk on the prey. The ‘ecology of fear’ concept states that prey modify their behaviour by striking a balance for their need to forage against their need to avoid predators (Brown, Laundre & Gurung, 1999). Consequently, this trade-off may result in the avoidance of food-rich habitat patches, either spatially or temporally, which remain unoccupied by prey species if these patches also have significantly higher predation risks. Such risk has been shown to affect physiological and demographic patterns of elk, Cervus elaphus preyed on by grey wolves, Canis lupus (Creel et al., 2007) and spatial patterns of bighorn sheep, Ovis canadensis, avoiding open habitats that provide greater visibility for pumas, Puma concolor (Altendorf et al., 2001). As no studies exist of Malayan tapir temporal patterns in areas without tiger, we speculate that the tapir's strong nocturnal activity patterns is advantageous for avoiding its only predator in KSNP the tiger. In comparison with the tapir, the pig-tailed macaque had an overall overlap coefficient almost identical to that of tapir, but in contrast was strongly diurnal, with significant activity occurring in the middle of the day, which is also the hottest time of the day. During this period, tiger activity was low, presumably because the species was resting. The limited data for wild pig suggest that it is also strongly diurnal (Laidlaw & Shaharuddin, 1998). Finally, there were several other putative prey species recorded in KSNP, argus pheasant Argusianus argus, mouse deer Tragulus spp., porcupine Hystrix brachyura and bearded pig Sus barbatus that may have influenced tiger temporal patterns. However, these were not included in the study as they were not considered to represent principal prey species because of their smaller body size (Karanth & Sunquist, 1995; O'Brien et al., 2003) or, in the case of the migratory bearded pig, an irregular food source. Ideally, tiger scat samples would have been collected for a dietary analysis of prey species composition, but scats are notoriously difficult to collect in tropical forests, because of low tiger population densities and high scat decay rates, and none were encountered during our field surveys. However, in the absence of difficult-to-collect dietary data, it is also valuable to demonstrate the temporal relationships, as conducted in this study, to provide new and much needed insights into Sumatran tiger–prey interactions. The methodology used here has wide application, especially for future statistical studies of predator–prey interactions or interspecific species competition.
From Tiger biologist and expert - Dr John Seidenstecker
The small Javan tiger ( weighing just 100 kg ) regularly preyed on wild Banteng and killed huge Banteng bulls weighing more than 800 kg:  *This image is copyright of its original author  *This image is copyright of its original author https://archive.org/details/tigers00seid...ker+tigers  *This image is copyright of its original author http://citeseerx.ist.psu.edu/viewdoc/dow...1&type=pdf
From leading tiger expert & scientist - Ullas Karanth
Tigers routinely kill adult male gaur that exceed the size of sub-adult male Rhinos:  *This image is copyright of its original author https://books.google.co.uk/books?id=8vco...ur&f=false Tigers routinely kill adult gaur, five times their own weight:  *This image is copyright of its original author https://books.google.co.uk/books?id=W6ks...ht&f=false Tigers do take the largest suids, bovids and cervids. In Kanha National Park in central India, Schaller (1967) found that tigers killed gaur and swamp deer when they were part of the ungulate assemblages. Karanth (1988) reported heavy predation on solitary adult gaur in Nagarahole National Park in southern India:  *This image is copyright of its original author http://citeseerx.ist.psu.edu/viewdoc/dow...1&type=pdf From George Schaller One of the largest Bull gaurs ever seen at Kanha was killed by a tiger: 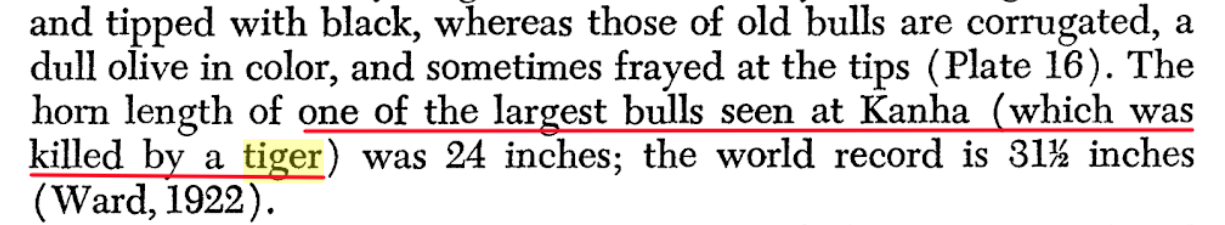 *This image is copyright of its original author https://books.google.co.uk/books?id=KOVG...er&f=false Tigress killed a huge 2000-pound Bull gaur:  *This image is copyright of its original author https://archive.org/details/naturalistot...her+beasts Gaurs and water buffalos weighing over a ton have been killed by tigers, weighing about a sixth as much:  *This image is copyright of its original author https://www.cms.int/sites/default/files/...only_0.pdf |





| Users browsing this thread: |
| 6 Guest(s) |