WildFact
›
Information Section
›
Extinct Animals
›
Pleistocene Big Cats

 Giant cheetah-Acinonyx Pardinensis
Giant cheetah-Acinonyx Pardinensis
Giant cheetah-Acinonyx Pardinensis
|
Giant cheetah-Acinonyx Pardinensis
|
|
Reconstructions by Roman Utchyel
-738x591.jpg) *This image is copyright of its original author  *This image is copyright of its original author  *This image is copyright of its original author
Giant cheetah weight estimates from Evolution of community composition in several carnivore palaeoguilds from the European Pleistocene: the role of interspecific competition, Garcia and Virgos 2007
 *This image is copyright of its original author Minimum weight:105 kg Maximum weight:140 kg Total average: 118 kg (n=3)
Measurements (mm) of canine and upper teeth of Giant cheetah from Pantalla
 *This image is copyright of its original author Measurements mm of mandible and lower teeth of giant cheetah from Pantalla  *This image is copyright of its original author From Acinonyx pardinensis Carnivora Felidae from the Early Pleistocene of Pantalla Italy predatory behavior and ecological role of the Giant Plio Pleistocene cheetah
Synchrotron radiation reveals the identity of the large felid from Monte Argentario (Early Pleistocene, Italy)
Abstract We describe here a partial skull with associated mandible of a large felid from Monte Argentario, Italy (Early Pleistocene; ~1.5 million years). Propagation x-ray phase-contrast synchrotron microtomography of the specimen, still partially embedded in the rock matrix, allows ascribing it reliably to Acinonyx pardinensis, one of the most intriguing extinct carnivorans of the Old World Plio-Pleistocene. The analysis of images and 3D models obtained through synchrotron microtomography – here applied for the first time on a Plio-Pleistocene carnivoran – reveals a mosaic of cheetah-like and Panthera-like features, with the latter justifying previous attributions of the fossil to the extinct Eurasian jaguar Panthera gombaszoegensis. Similarly, we reassign to A. pardinensis some other Italian materials previously referred to P. gombaszoegensis (sites of Pietrafitta and Ellera di Corciano). The recognition of Panthera-like characters in A. pardinensis leads to reconsidering the ecological role of this species, whose hunting strategy was likely to be different from those of the living cheetah. Furthermore, we hypothesise that the high intraspecific variation in body size in A. pardinensis can be the result of sexual dimorphism, as observed today in all large-sized felids. Head appearance of Acinonyx pardinensis. The reconstruction is based on the complete skull from Pantalla6 and is updated with the information on the craniodental anatomy of A. pardinensis achieved from the Monte Argentario specimen. Artwork by D. A. Iurino.  *This image is copyright of its original author Acinonyx pardinensis from Monte Argentario. Specimen ArgBsc1 in right lateral (a), rostral (b), and left lateral (c) views. Scale bar: 50 mm.  *This image is copyright of its original author Results External observations The specimen (ArgBsc1) is represented by the rostral portion of the skull with associated mandible of a large felid (Fig. 2, Table 1). The neurocranium is almost completely missing, with only a small portion of the left frontal bone being visible. The splanchnocranium is better preserved on the left side, from which it looks rather shortened and dorsoventrally expanded. The zygomatic arches are lacking, as well as the orbits, with the exception of the dorsal margin of the left one, which is delimited by the postorbital process of the frontal. In rostral view, the nasal apertures look broad though slightly distorted, with the median part of the nasal bones being crushed into the nasal cavities. The mandible preserves the complete right corpus articulated with the anterior portion of the left one, broken at the level of the distal margin of the p4. Two mental foramina are visible on both sides, located at the same height immediately behind the distal margin of the lower canine and a few centimetres behind, respectively. Both the ascending rami are lost. The teeth are virtually unworn and in variable state of conservation. Only the roots of the right I3 and i3 are visible in rostral view. The left canines are missing, but the distoventral portion of the lower canine root is still preserved, delimiting a wide pulp cavity. Conversely, the canines on the right side are almost intact and stand out to be robust and elongated. A short distance separates the upper canine from the P3. The latter is broken on the right side, but complete on the left. It is characterised by a high, vertical, and pointed paracone. The right P4 is a large and robust tooth. In labial view, it shows, from front to back, a strong parastyle, a high and vertically-oriented paracone and a slightly shorter metacone. The root of the small M1 is visible distally to the right carnassial. With the exception of the canine, the other lower teeth are impossible to describe because they are covered by the upper teeth or the embedding matrix. The latter is a highly consolidated red-rust coloured claystone very hard to remove mechanically without the risk of damaging the fossil. This led to the need to examine the specimen through a highly performing x-ray image analysis technique. Discussion The identification of the specimen (ArgBsc1) based solely on external observation is hard, since the enclosing in the rock matrix makes very difficult the analysis of diagnostic characters, especially in the teeth. From the outside, the skull shows a series of intermediate Acinonyx-like and Panthera-like characters. The first include the high and short muzzle, the straight ventral margin and the right rostroventral angle of the mandible. The second include the relatively narrow frontals and especially the massive canines, mainly the upper ones, housed in prominent canine eminences. In particular, the latter characters might have led to the past erroneous attribution of the specimen to P. gombaszoegensis18, whose stratigraphic occurrence partially overlaps that of A. pardinensis. This may have been due also to the limited past knowledge of this taxon’s anatomy, which has increased significantly thanks to new discoveries in recent decades. In this regard, it is worth mentioning two other cases of misleading identification involving the same taxa. The first concerns Pietrafitta, a central Italian site with an age comparable to that of Monte Argentario25, the second the slightly older locality of Ellera di Corciano26. The scarce felid material from the two sites were referred in the past to P. gombaszoegensis27,28,29,30 and cf. P. gombaszoegensis31,32, respectively. The fossils are here reassigned to A. pardinensis (Supplementary Note 1 and Supplementary Figs S1–S2). The study of images obtained through the SR-µCT survey resulted in many additional information for the definition of the taxonomic status of the Monte Argentario skull. The following craniodental features identified on the 3D models of ArgBsc1 are consistently found in the other specimens attributed to A. pardinensis6,12,33 and differ from pantherine felids:
Similarly, most of the above characters allow to exclude that the Monte Argentario specimen can be referred to the late Pliocene-Early Pleistocene puma-like felid Puma pardoides34,35. In fact, although the latter species shows some similarities with A. pardinensis (e.g., lateral enlargement of the frontals caudally to the zygomatic processes and relatively shortened muzzle)6,33, differences between them are remarkable (e.g., Pu. pardoides has labiolingually enlarged P3, P4 with strong Panthera-like protocone, p3 significantly lower than p4 in labial view both showing more massive protoconid)5,6. Moreover, the overall size of Pu. pardoides is significantly smaller than that of A. pardinensis (the upper carnassial of Pu. pardoides is about 22 mm in average length5, while that of A. pardinensis is about 30 mm6). In the light of the above features, the felid from Monte Argentario is here referred to A. pardinensis. However, the occurrence of Panthera-like characters in the skull is noteworthy. Among them, the presence of stout and strong canines stands out. The linear dimensions (length and breadth) of these teeth fall in the uppermost part of the A. pardinensis range and, at least for the upper canine, are close to the lower values recorded for medium-sized pantherines like P. gombaszoegensis (Supplementary Tables S1–S2 and Supplementary Fig. S3). Similarly, the comparative analysis of the palatal sutures (clearly visible in ArgBsc1 for the first time in A. pardinensis) evidences further affinities with pantherine cats. In particular, the indented suture trend observed in the Monte Argentario skull recalls more the extant leopard and snow leopard than the cheetah and puma (Supplementary Fig. S4). Previous studies6,10,12 already highlighted that A. pardinensis retains Panthera-like, primitive morphological characters, but most of them relate to the neurocranium (e.g., the relatively elongated braincase, with high sagittal and nuchal crests and—as also visible in ArgBsc1—narrow frontals between the postorbital processes). The specimen from Monte Argentario shows some significant pantherine-like characters also in the splanchnocranium (i.e., teeth and palatal sutures). Nevertheless, at least as regards to canine dimensions, our results should be placed in the context of the overall A. pardinensis intraspecific variation. As in part predictable by the long-lasting stratigraphic distribution (from late Pliocene to Middle Pleistocene) and the wide geographical distribution (from Morocco to China), A. pardinensis exhibits a considerable variation in size, with body mass estimated (for adult individuals) from about 60 to 120 kg (Supplementary Table S3 and Supplementary Fig. S4). This variation led some scholars15,33,36 to consider A. pardinensis as a broad ‘macrospecies’ subdivided into several subspecies6: A. p. arvernensis from Les Étouaires (late Pliocene) and Tuozidong (early Early Pleistocene); A. p. aicha from North Africa (earliest Pleistocene); A. p. pardinensis from a number of European to central Asian sites (Early Pleistocene; Middle-Late Villafranchian LMA); A. p. linxiaensis (=‘Sivapanthera’ linxiaensis)10 from northern China (early Early Pleistocene); A. p. pleistocaenicus from Untermassfeld (late Early Pleistocene; Epivillafranchian stage)37 and China (=‘Sivapanthera’ pleistocaenica)11 (Early Pleistocene; Villafranchian LMA); A. p. intermedius from Hundsheim and Mosbach (Middle Pleistocene; Galerian LMA). However, the validity of this taxonomic model is questioned by the lack of clear morphological or biometrical differences between the different subspecies. The available estimated body masses for A. pardinensis suggest that the variation found is independent of the age and the geographical location of the finds (Supplementary Fig. S5). An emblematic case is represented by the earliest (i.e., Early Villafranchian LMA) European specimens attributed to this taxon, namely those of Villafranca d’Asti (Supplementary Fig. S6), Villarroya, and Les Étouaires, for which the predicted body masses are 70, 87, and 121 kg, respectively. Unfortunately, the existing data are not sufficient to understand if the intraspecific variation of A. pardinensis could be linked to ecogeographical dynamics and/or sexual dimorphism. However, actualistic comparisons with extant large-sized felids allow to hypothesise that the latter factor can be called into question for A. pardinensis. Sexual dimorphism in body mass occurs in all living felids, and is particularly marked in wide-range large-sized species. For example, the average weight difference between males and females is about 60% in the jaguar, 80% in the leopard, 85% in the lion and the tiger, and even 90% in the puma, while it is much smaller (~15%) in the cheetah38. We obtain interesting information if we try to free the interpretations on A. pardinensis from the stratigraphic influence, that is if we consider the few available sites for which it is possible to estimate body mass for more than one individual (which we can therefore hypothesise as being coeval). The difference between the lowest and highest predicted body mass is about 31% at Ahl al Oughlam, 33% at Saint Vallier, and 72% at Longdan. This suggests that sexual dimorphism may have played an important role in the intraspecific variation of A. pardinensis, as hypothesised by Petrucci et al.39 based on postcranial material from Pirro Nord. Although out of the scope of this work, the same can be said for the Eurasian jaguar P. gombaszoegensis, whose predicted body masses span from about 65 to 180 kg in all its chronological (about 1.9–0.3 Ma) and geographic (Europe and Western Asia) range, with peaks of variation of ~80% and even 90% for local samples such as those from the type locality of Gombaszög and Untermassfeld, respectively (Supplementary Table S4). Acinonyx pardinensis and P. gombaszoegensis co-occurred in Europe for about 1.4 Ma in the Early and Middle Pleistocene, and are reported together in at least seven sites (Olivola, Dmanisi, Upper Valdarno, Pirro Nord, Untermassfeld, Le Vallonnet, and Mosbach, in decreasing stratigraphical order), often in association with sabretooth cats like Megantereon and/or Homotherium6,39,40. This extraordinary concentration of large felids is unequalled in modern ecosystems and, moreover, must be included in the context of the rich diversity of the Late Villafranchian/Epivillafranchian large carnivore guilds, which included, among others, also hyaenids and canids41. Interspecific competition for prey must have been strong in such palaeoecosystems and subject to ecological mitigation strategies. These could have included temporal niche displacement (e.g., diurnal/nocturnal hunting) and/or different spatial distribution within the ecosystem6 (for example, in Venta Micena, geochemical data suggest that Megantereon and Panthera preferred forested areas whereas Homotherium and Lycaon hunted in more open habitats)42. The recognition of pantherine-like characters in the Monte Argentario felid—as well as in other A. pardinensis samples from other localities—must lead to reconsidering the ecological role of this species, whose choice of prey and hunting strategy was likely to be different from those of the living cheetah6,16. Furthermore, we hope that the approach followed in this work, i.e., the use of high-resolution µCT techniques, may also be followed in the future for similar cases, in order to extract as much information as possible from fossil remains of Plio-Pleistocene vertebrates.
Hemmer, H., Kahlke, R.-D., and Vekua, A. K. (2011). The cheetah Acinonyx pardinensis (Croizet et Jobert, 1828) s.l. at the hominin site of Dmanisi (Georgia) - A potential prime meat supplier in Early Pleistocenen ecosystems. Quaternary Science Reviews 30: 2703-2714.
Abstract: The fossil site of Dmanisi (southern Georgia) has yielded a significant amount of hominin remains dated to around 1.8 Ma, in addition to a rich contemporaneous faunal record. Based on topographic information combined with an updated list of the vertebrate faunal assemblage, the corresponding palaeo-landscape has been reconstructed. Over a distance of some kilometres the landscape pattern changed from that of a forested valley floor, to tree savannah and open grasslands, thus providing typical habitats for carnivores hunting in open spaces. Morphological analysis of the elements from a nearly complete cat's foreleg reveals the existence of a large and stoutly built cheetah, Acinonyx pardinensis (Croizet et Jobert, 1828) s.l., in the Dmanisi faunal assemblage. Body mass estimations based on the humerus and metacarpals point to a cat of around 100 kg. The amount of pure meat and associated leftovers produced by the cheetah's hunting activity available for other consumers has been estimated. Within Early Pleistocene ecosystems, the cheetah must be considered as a potential fresh prime meat supplier, above that of any other felid. 1. Introduction In addition to a rich, contemporaneous vertebrate faunal record, the fossil site of Dmanisi in southern Georgia, 85 km SW of Tbilisi (Fig. 1), has yielded a large amount of hominin cranial, dental and postcranial elements (Gabunia et al., 2000; Vekua et al., 2002; Rightmire et al., 2006; Lordkipanidze et al., 2007; Martinón-Torres et al., 2008). By placing the Dmanisi faunal assemblage at around the Olduvai/Matuyama palaeomagnetic boundary (Lordkipanidze et al., 2007), the corresponding hominin remains appear to be the stratigraphically oldest indication of Homo outside of Africa. These remains demonstrate the advance of humans from Africa via the Levantine Corridor and into Transcaucasia at the onset of the Late Villafranchian, around 1.8 Ma. Before the discovery of an almost complete set of skeletal elements of a large cat’s foreleg at Dmanisi, two machairodonts, Homotherium crenatidens (Weithofer, 1889) and Megantereon cultridens (Cuvier, 1824), were the only felids known from the mammal assemblage of the site (Vekua, 1995). After the subsequent discovery of a mandible of an Eurasian jaguar (recently named Panthera onca georgica Hemmer, Kahlke et Vekua, 2010), the limb bones in question were attributed to this species, as it was impossible to relate them morphologically to a machairodontine cat (Vekua, 2001). In addition to the three cat species so far recorded at Dmanisi, the guild of western Palaearctic large felids around 1.8 Ma comprised the Eurasian puma, Puma pardoides (Owen, 1846) (Hemmer, 2004), and the large cheetah, Acinonyx pardinensis (Croizet et Jobert, 1828). In view of the existence of many open landscape dwellers in the herbivore record, and a correspondingly reconstructed landscape pattern, the presence of, at least, the latter of these felid species was expected in the Dmanisi palaeo-ecosystem. A re-examination of the above-mentioned postcranial felid material from Dmanisi did, in fact, result in the discovery of a cheetah. Based on the comparative description and evaluation of the most diagnostic skeletal elements of its foreleg, the taxonomic affiliation of this new element of the Dmanisi fauna is discussed here, along with a review of the relevant material from Eurasian Plio- and Early Pleistocene localities. By applying different prediction equations, a body mass estimation of this large predator has been calculated. Combined with its inferred hunting success, this new data provides a clearer understanding of the role of cheetahs within Early Pleistocene ecosystems. 2. Material and methods Based on a number of studies on different faunal elements, in addition to new observations, we present here an updated vertebrate faunal list for the site of Dmanisi. The attribution of the majority of recorded species to corresponding biotopes, through consideration of the given geological and topographic situation, has enabled a reconstruction of the Dmanisi palaeo-landscape pattern during the period of site formation to be put forward. The studied felid material was found during the 1997 field campaign (square 63/59) and is stored in the collection of the Georgian National Museum Tbilisi (inventory numbers: Scapula D1999, Humerus D2000, Radius and Ulna D2001, Metacarpals IIeV D2002aed, phalanx I-III D2003e2005). The most diagnostic elements are presented in this study. Some unpublished Late Plioand Early Pleistocene humerus fragments from Villafranca d’Asti (Italy) (NHMB V.J.133) and St. Vallier (France) (NHMB St.V.265, 266) were studied in the Basel Natural History Museum. Measuring was done by H.H. using classic dimensions. The set of specific measurements published by Schütt (1970) was used for the distal humerus diagnosis. To estimate the body mass of the newly discovered Dmanisi cat, a set of prediction equations was calculated based on the metacarpal shaft minimum diameters. The following data were used to this end: body masses were taken as species means, or as male and female means, respectively (if the available metacarpal data were clearly assignable to a sex), based on Nowell and Jackson (1996) and Piechocki (1986). These data were rounded as follows: to the nearest ½ kg for means lower than 5 kg; to the nearest 1 kg for cats weighing more than 5 but less than 20 kg; to the nearest 5 kg for the range between 20 and 50 kg; to the nearest 10 kg for specimens weighing more than 50 but less than 200 kg (cf. Hemmer, 2004). Metacarpal shaft dimensions were collected from Del Campana (1916, 1954), Hooijer (1947), Thenius (1954), Kurtén (1965, 1978), Bjork (1970), Clot and Besson (1974), Kurtén and Crusafont-Pairó (1977), Van Valkenburgh et al. (1990), Morgan and Seymour (1997), García García (2003, p. 100), and Morales et al. (2003). Dimensions of Felis catus were taken by one of the authors (H.H.). Used alongside behavioural data observed from extant cheetahs, both the updated palaeoenvironmental data from Dmanisi and the morphometric analysis of the discussed felid remains, have shed light on the palaeoecological role played by large sized cheetahs in Early Pleistocene ecosystems of the western Palaearctic. 3. Early Pleistocene fauna and palaeo-landscape reconstruction of the Dmanisi site 3.1. Vertebrate faunal assemblage Apart from the hominin record, the Dmanisi fossil vertebrate assemblage has so far produced the remains of 50 taxa of amphibians and reptiles (8), birds (3) and mammals (39): Bufo viridis Laurenti, 1768, Anura gen. et sp. indet., Testudo graeca Linnaeus, 1758, Lacerta ex. gr. viridis Laurenti, 1768, Sauria gen. et sp. indet., cf. Elaphe quatuorlineata (Lacépède, 1789), Colubines gen. et sp. indet., Serpentes gen. et sp. indet., Struthio dmanisensis Burchak-Abramovich et Vekua, 1990, Gallus dmanisiensis Bendukidze et Burchak, 2001, Strix gigas Bendukidze et Burchak, 2006, Ochotona ex gr. Ochotona lagreli minor Bohlin, 1942, Hypolagus cf. brachygnathus Kormos, 1934, Marmota sp., Cricetus sp., Mimomys tornensis Jánossy et van der Meulen, 1975, Mimomys ostramocensis Jánossy et van der Meulen, 1975, Gerbillus sp., Parameriones obeidiensis (Haas, 1966), Apodemus dominans Kretzoi, 1959, Hystrix sp., Beremendia fissidens (Petényi, 1864), Lynx issiodorensis (Croizet et Jobert, 1828), Panthera onca georgica Hemmer, Kahlke et Vekua 2010, Megantereon cultridens (Cuvier, 1824): pending relation to the African Megantereon whitei (Broom, 1937), Homotherium crenatidens (Weithofer, 1889), Pachycrocuta perrieri (Croizet et Jobert, 1828), Pachycrocuta sp., Canis etruscus Forsyth Major, 1877, Vulpes alopecoides Forsyth Major, 1877, Ursus etruscus Cuvier, 1812, Ursus sp., Meles sp., Martes sp., Insectivora gen. et sp. indet., Dama (¼ Pseudodama ¼ Cervus s.l.) cf. nestii Forsyth Major, 1879, Cervus abesalomi Kahlke, 2001 (¼ Cervus perrieri Croizet et Jobert, 1828 sensu Vekua, 1995), Eucladoceros aff. tegulensis (Dubois, 1904) [ ¼ E. aff. senezensis (Depéret, 1910)], Palaeotragus sp., Bison (Eobison) georgicus (Burchak-Abramovich et Vekua, 1994), Pontoceros sp., Fig. 1. Geographic position of the Early Pleistocene hominin site of Dmanisi (Georgia). 2704 H. Hemmer et al. / Quaternary Science Reviews 30 (2011) 2703e2714 Antilopini gen. et sp. indet., Gallogoral meneghinii sickenbergii Kostopoulos, 1996, Soergelia cf. minor, Ovibovini gen. et sp. indet, Capra dalii Bukhsianidze et Vekua 2006, Equus stenonis Cocchi, 1867, Equus sp. aff. altidens von Reichenau, 1915, Stephanorhinus etruscus (Falconer, 1859), Mammuthus meridionalis meridionalis (Nesti, 1825) (compiled under consideration of the contributions by Vekua and Gabunia, 1989; Gabunia and Vekua, 1993; Vekua, 1995; Bukhsianidze, 2005, 2006; Bukhsianidze and Vekua, 2006; Lordkipanidze et al., 2007; Palmqvist et al., 2007; Vekua et al., 2008; Furió et al., 2010; Hemmer et al., 2010). Whereas the Late Pliocene faunas of Transcaucasia show remarkable affinities to Africa, as recorded from the c. 3 Ma old site of Kvabebi in eastern Georgia (Vekua, 1972; updated faunal lists in Hemmer et al., 2004; Agustí et al., 2009), the northern dispersal of African species into Eurasia was more limited during the Middle (c. 2.6e1.8 Ma) and Late Villafranchian (c. 1.8e1.2 Ma) (Kahlke et al., 2011). The earliest Late Villafranchian fauna of Dmanisi is of, predominantly, Eurasian character. Several of the faunal elements recorded at Dmanisi are also common in the Middle Villafranchian contexts of Western Asia and Europe: Gallogoral meneghinii sickenbergii, Eucladoceros aff. tegulensis, Palaeotragus sp., Mammuthus meridionalis (typical form) and Pachycrocuta perrieri. More modern forms comprise Bison (Eobison) georgicus, Pontoceros sp., Dama cf. nestii, and the elaphine Cervus abesalomi. 3.2. Palaeo-landscape reconstruction after the vertebrate record The fossil bone concentrations at the site of Dmanisi were accumulated in a broad river valley whose floor consisted of basalts and volcanoclastic alluvials, near to the confluence of two successively incising river branches (Dzaparidze et al., 1989). However, the Dmanisi taphocoenosis does not only include species which inhabited valley floodplains. Lake and river landscapes with their wet biotopes and open water bodies attract many species and are regularly frequented for drinking, and in some cases for food resources, by species roaming in both the immediate surroundings, as well as in the broader environs of the valley. Qualitative analyses can be performed on indicator species, which enable the different types of habitats present in the Dmanisi region during the site formation to be described. In order to quantify the share held by each of the different type of habitats, an unbiased quantitative representation of all faunal elements is required. As far as the taphonomic evidence is understood (Tappen et al., 2007), this prerequisite is lacking at Dmanisi. Nevertheless, the available palaeozoological record provides a detailed picture of the shape of the landscape whilst the Dmanisi fossil site was evolving. The immediate vicinity of (at least seasonal) fluviolimnic water bodies is indicated by the geographic position of the site in a flat river valley, now forming a promontory above the confluence of the Mashavera and Pinezaouri Rivers. In the more stable areas of the palaeo-valley, the presence of a gallery forest is supported by the numerous cervid remains recovered from the site. In addition to the frequently recorded Eucladoceros aff. tegulensis, Cervus abesalomi must also be assigned to wooded biotopes. However, the regular occurrence of Dama cf. nestii in more open landscapes cannot be excluded (cf. Hemmer, 2001). Bison (Eobison) georgicus might have been predominantly a woodland inhabitant, as were all of the, so far identified, members of the subgenus Eobison (Kahlke, 1999, p. 109). Jaguars (Panthera onca georgica) roamed in different climatic and vegetation zones of Eurasia from the latest Middle Villafranchian onwards up to the early Middle Pleistocene. These cats preferred areas where gallery forests provided a suitable spectrum and quantity of prey (Hemmer et al., 2003). Dirktooth cats (Megantereon) chiefly hunted in forest habitats, as has been shown by both comparative morphology as well as biogeochemical data (Hemmer, 2001; Palmqvist et al., 2007). The records of Strix, Meles and Apodemus dominans also support the idea of gallery forest biotopes in the vicinity of the Dmanisi site. Stephanorhinus etruscus and Mammuthus meridionalis meridionalis lived in open wooded areas as well as in savannah like landscapes (Kahlke, 2006). Garutt and Bajguseva (1981) were able to demonstrate that the high and relatively short stature of the Southern Elephants allowed them to forage on leaves, branches and bark. It is likely that their dietary spectrum showed a marked seasonal variation. Whereas fresh steppe vegetation may have been preferred in spring and at the beginning of summer, arboreal forage probably became more important later in the year. The woodland steppes and tree savannahs would also have been a natural habitat for Palaeotragus (Godina, 1979) and so, away from the gallery forest, landscapeswithminor tree vegetation and open areas have to be assumed. The overall picture of the Dmanisi fauna is characterised by numerous species typically associated with open environments. In addition to Struthio dmanisensis, Hypolagus cf. brachygnathus, Cricetus, Gerbillus, Parameriones obeidiensis and the equids, as well as members of the Antilopini group, all indicate the presence of savannah to steppe conditions away from the river valley. The Lacerta viridis prefers sunny areas near to dense shrub vegetation, and Bufo viridis is able to live in open steppe biotopes. Furthermore, the presence of certain carnivore species in the fossil record, e.g. Vulpes alopecoides with its more or less developed affinity to open steppe biotopes (Kahlke, 1999), also support the idea of a predominantly open environment for the Dmanisi palaeo-landscape. If the recorded Pachycrocuta perrieri is, indeed, a brown hyaena, as discussed (Turner, 1990), its habitat preferences would, most likely, have been similar to that of the extant south African Hyaena brunnea, which roams predominantly in dry savannah, steppe and semi-desert landscapes. Meanwhile, the presence of Testudo graeca and Hystrix are indicative of temperate Mediterranean climatic parameters. The presence of the gallinaceous bird, Gallus dmanisiensis, suggests the existence of a plain at the foothills of a mountainous landscape, most probably with bush cover (Bendukidze and Burchak, 2001). The occurrence of Soergelia cf. minor seems to be an indicator of a more pronounced palaeo-relief or possibly even mountainous conditions. Picas, like the recorded Ochotona ex gr. Ochotona lagreli minor, are found in areas of broken rock and talus that are surrounded by suitable vegetation. They are most commonly seen at the interface between meadow habitat and open rocky terrain. Typical inhabitants of rocky mountain areas are the recorded ibex Capra dalii, which would have avoided open plains, as well as Gallogoral meneghinii sickenbergii, which has been characterised, using ecological analogies, to extant Nemorhaedus and Capricornis (Guérin, 1965). Some of the recorded carnivore species, i.e., Lynx issiodorensis, Homotherium crenatidens, Canis etruscus and Ursus etruscus, did not require specialised habitats, but could adapt well to any of the landscape components described above. The combination of topographic and vertebrate palaeontological information provides strong evidence of the existence of a fairly differentiated landscape pattern around the site of Dmanisi. The flat river valley, c. 1,300 m above sea-level, would have contained open water bodies and areas of wet ground throughout the year. Other parts of the valley floodplain were covered by gallery forests. Due to the lithologic features of their volcanic subsoil, the flanking slopes of the river valley would have been significantly dryer and poorer in tree cover. They would have been dominated by a lower diversity of dense shrub vegetation lying adjacent to more open areas. On the south to southeast facing exposed slopes dry meadows would have developed. Extended grasslands characterised the site vicinity in the adjacent higher ground outside of the valley. In addition to dry H. Hemmer et al. / Quaternary Science Reviews 30 (2011) 2703e2714 2705 savannahs, even semi-desert like areas could have existed in the Dmanisi area. Over distances of just a few kilometres, cuesta scarps of basaltic lavas (today’s relative height 75-150 m) would have led to the formation of broken rocks, taluses and rocky terrain, in which ibexes and gorals would have found suitable habitats. These animals were probably related to corresponding populations in the Dzavacheti mountain chain (up to 3000 m above sea-level) some 20 km west of the site. The palaeo-landscape of the Dmanisi area showed remarkable gradients in its relief as well as in its humidity and vegetation character. Over a distance of some kilometres the landscape pattern appears to have changed dramatically, from that of a forested valley floor and slopes with shrub vegetation, to tree savannah and open grasslands, and up to semi-arid areas and rocky terrains. Whereas the ecological niche of deer and bison hunters, in the more forested areas, was occupied by the jaguar and the dirktooth, the more ubiquitous elephant and rhino populations were most probably exploited by the large sabretooth. However, the presence of a specialized hunter which targeted the antelope to horse sized game, in the predominantly dryer and more open areas of the site’s vicinity, had, until now, been absent from the fossil record. 4. Morphometric analysis of the Dmanisi Acinonyx remains 4.1. Brachial index The availability of associated and complete long bones of an anterior limb has enabled the first substantial attempt using the brachial (humeroradial) index (radius length/functional humerus length 100), as calculated for large felids, e.g. by Vereshchagin (1971) or by Gonyea (1976). An index value of 99.5 was obtained for the Dmanisi cat. This value differs from the means of both Panthera onca (86.8 2.08) and Panthera pardus (90.5 1.46) by over 6 standard deviations, and from Panthera tigris (89.8 0.89) by > 10 s. These extant pantherine species must, therefore, be excluded as candidates in the identification of the Dmanisi find. The humeroradial ratio of the Dmanisi cat fits the distribution recorded for extant Panthera leo (98.3 2.22) (means and standard deviations by Gonyea, 1976), as well as the variability of Acinonyx jubatus. Two data sets were published for this species, 103.3 1.45 (Gonyea, 1976) and 96.7e10 4.2 (Vereshchagin, 1971). In view of low standard deviations found in extant large felids, the brachial index also qualifies as a useful tool in the definite discrimination between the Dmanisi cat and the Pliocene-Pleistocene machairodont species Homotherium crenatidens (89.5), Megantereon cultridens (81.6), and Dinofelis barlowi (80.5 0.50) (data by Marean, 1989; see also Werdelin and Lewis, 2001). Furthermore, the brachial index of the Dmanisi cat excludes identification with the Blancan/Irvingtonian North American cheetah-like, but puma-related (Barnett et al., 2005), cat, Miracinonyx inexpectatus (91.1 respectively around 90; Van Valkenburgh et al., 1990). In contrast, the associated long bones of a large European cheetah from Étouaires (France), dated to the end of the Lower Villafranchian (MN 16), have a humeroradial ratio which matches the value of the Dmanisi individual. The humerus length of the Étouaires specimen has been given as 267 mm and the radius length as 268 mm by Croizet and Jobert (1828). Van Valkenburgh et al. (1990) measured the radius length (cast) as 266 mm (a humerus length of 260 mm cited by these authors in their Table 3 obviously is the result of a print error, as Fig. 6 of their paper shows nearly the same humerus as radius lengths). Savage (1960) gave the radius length, using the same cast as Van Valkenburgh et al. (1990), as c. 269 mm. In view of these different values, Croizet and Jobert’s measurements on the originals have been used in this study, resulting in a brachial index of 100.4. The Early Pleistocene cat from Dmanisi discussed here largely predates the first appearance of the lion in Eurasia during the early Middle Pleistocene. Moreover, the Eurasian cave lion’s radius is relatively shorter (brachial index 91.5 after Vereshchagin, 1971) than that of the recent African lion (see above). Consequently, the brachial index of the Dmanisi foreleg also excludes a Panthera leo diagnosis. Using the brachial index diagnostic tool, therefore, the genus Acinonyx is considered to be the only suitable candidate adequately fitting the Dmanisi measurements. 4.2. Foreleg skeletal elements 4.2.1. Humerus The humerus is strikingly slender, especially the distal region, when compared to other large-sized felids (Fig. 2, Table 1). A jaguar’s humerus of the same length would have a 25% larger distal transverse diameter than that recorded in the Dmanisi fossil (log distTD ¼ 1.16 log L e 0.95, n ¼ 6, r ¼ 0.978; basic data for this allometric equation: 1 extant þ 5 Pleistocene American jaguars; Kurtén, 1965, 1973). The Dmanisi measurement falls outside the lower 95% confidence limit of jaguars (width to length scatter), as do extant cheetahs and the Acinonyx humerus of Étouaires. In contrast, the Dmanisi humerus compares well with the humerus of the late Early Pleistocene (Irvingtonian) Hamilton Cave Miracinonyx inexpectatus skeleton. In total, the Miracinonyx/Puma-group (data by Van Valkenburgh et al., 1990) ranges just around the lower 95% confidence limit of Panthera onca, but above the cheetahs. The distal part of a cheetah humerus is characterised by a weak entepicondylus, resulting in an unusually large trochlear transverse diameter when related to the distal width. The trochlea is also comparatively high and thick. Its upper margin to the fossa olecrani runs nearly horizontal; the fossa olecrani itself is formed like a high pointed arch. The posterior entrance of the foramen entepicondyloideum is positioned high at the shaft (Schütt, 1970). Indices, which allow morphometric expression of these specific characters, demonstrate that the humerus of the Dmanisi cat significantly differs from that of Panthera humeri, but is affiliated to those of Acinonyx. The relative width of the trochlea distinguishes it from those of the Miracinonyx/Puma-group (Table 2). In conclusion (and despite of some similarities to the Nearctic Miracinonyx inexpectatus), the Dmanisi humerus discussed here has been attributed to the genus Acinonyx. This element represents the largest humerus of an extinct or extant cheetah on record, and is clearly stouter than the insignificantly shorter humerus from Étouaires. The latter fossil is not presented in Table 2, as the published distal transverse diameter measurements are contradictory: 44 mm in Croizet and Jobert (1828; the humerus is listed here as belonging to a dirktooth cat), but c. 41.3 mm in Van Valkenburgh (1990). Moreover, the drawing given by Croizet and Jobert (l.c.) indicates some breakage at the entepicondylus. 4.2.2. Radius The distal transverse diameter to radius length scatter places the slender Dmanisi radius (Fig. 3) far outside the variability recorded for extinct and extant American Panthera onca, and even places it outside an extrapolation of the 99% confidence limits. Only specimens of Acinonyx and Miracinonyx range this far outside of the jaguars’ variation. Nearest to the range for the Dmanisi radius are those of the Étouaires specimen, which is even more slender than the Dmanisi element. Miracinonyx radii show comparable proportions, but are much shorter. Van Valkenburgh et al. (1990) highlighted some diagnostic characteristics which enable discrimination between the extant cheetah and the Puma/Miracinonyx-group. Proximally, the proximodistal length of the biceps tuberosity of Puma, Miracinonyx and Acinonyx pardinensis (Étouaires) has about twice the extension (near to 10% of the radius length) of that measured in Acinonyx jubatus (about 5%). The Dmanisi fossil fits the former group. Distally, in Puma and Miracinonyx the articular facet for the ulna is elliptical, with its long axis oriented perpendicular to the shaft, rather than in line with the shaft as seen in Old World cheetahs. In the Dmanisi specimen this facet appears to be nearly circular. Overall, the morphology of the Dmanisi radius matches closest to that of the Étouaires Acinonyx radius, though the latter looks slightly more advanced towards that of the extant cheetah. 4.2.3. Ulna The ulna of the Dmanisi cat (Fig. 4) is not completely preserved, a large part of the olecranon is missing. Thus, important measurements such as the greatest length and some olecranon diameters cannot be taken. Diagnostic characters of the distal part of the ulna, as comparatively shown by Van Valkenburgh et al. (1990) for Acinonyx jubatus, Miracinonyx inexpectatus and Miracinonyx trumani, in addition to Puma concolor, exclude the extant cheetah, puma and ancient M. inexpectatus - contemporaneous with the Dmanisi specimen - as possible candidates. The styloid processus of the latter resembles Acinonyx from Étouaires (Croizet and Jobert, 1828; Felis, Plate 1, Figs. 4 and 5). 4.2.4. Metacarpalia The metacarpals IIeV (Figs. 5e8) look very slender in comparison with the same elements from both extinct and extant jaguars. The most diagnostic MC V ratio diagram shows that the Dmanisi measurement even falls outside of the jaguar’s lower 99% confidence limit. In contrast, the Dmanisi values fit well with the allometric lines recorded for cheetah metacarpals from Pliocene through to extant specimens (Fig. 9). Measurements of Puma and Miracinonyx scatter slightly above the Acinonyx allometric lines. A safe morphometric discrimination of the Dmanisi metacarpals from those of the similar-sized Miracinonyx inexpectatus is not possible. 4.3. Body mass estimation The availability of a nearly complete leg of the Dmanisi cheetah permits an attempt to estimate its body mass. The most accurate results are most likely to be obtained using an approach based on the long bone shaft diameters, which are load-related and therefore provide a direct functional relationship to body mass (Hemmer, 2001, 2004, 2007a). So far, body mass prediction based on felid foreleg elements has been restricted to the smallest humerus shaft transversal diameter (Hemmer, 2001). In order to increase the likelihood of obtaining a reliable body mass estimation for the Dmanisi cat, new prediction equations were calculated for MC II to MC V shaft TDs, using various sets of published measurements (Table 3; see Section 2). We abstained from performing a similar calculation for the radius because the shaft transverse diameter is harder to define. Calculations with the new equation set for the Dmanisi cat resulted in the following body mass raw data: humerus 126 kg, MC II 92 kg, MC III 98 kg, MC IV 102 kg, MC V 76 kg, and mean 99 kg. The retained body mass estimate, rounded to the nearest 10 kg as recommended by Hemmer (2001, 2004) to avoid inappropriate precision, amounts to 100 kg. Overestimation of the body mass using the humerus diameter as well as underestimation by the MC V diameter requires interpretation. Generally, such differences in body mass estimation, based on different parts of a single skeleton, are not surprising, neither in felids nor in hominoids (Hemmer, 2001, 2007b). Morphometric analysis has shown that the Dmanisi humerus is substantially stouter than the (just insignificantly shorter) humerus of the Étouaires cheetah (see Section 4.2.1). This may indicate a particularly robust development of this section of the foreleg. Compared to the other elements of the front paw, the MC V is more lightly built in modern cheetahs than in other felids, thus suggesting a too small body mass when calculating with the general felid equation. For example, the MC V of an average modern cheetah used by Thenius (1954) for comparison with a metacarpal from the early Middle Pleistocene Hundsheim site (Austria) points to a body mass of 34 kg, although the mean body mass for extant African cheetahs is 43.2 kg, with a minimum to maximum span between 21 and 65 kg (Marker and Dickman, 2003, compiled from a total of 220 East, Southwest and South African cheetahs). The MC V of a small West African cheetah referred by Thenius (1954), results in an obvious underestimation of the body mass of 20 kg only. Consequently, body mass calculations based on the MC V of extant an 5. Discussion 5.1. Taxonomic affiliation of the Dmanisi cheetah The morphological analyses performed on the described Dmanisi cat remains (see Sections 4.1, 4.2.1e4.2.4) result in assignments to the genus Acinonyx. During the last four decades, Eurasian cheetahs of Late Pliocene and Early Pleistocene age (>3.0e0.8 Ma) were mostly attributed to one single species, A. pardinensis (Croizet et Jobert, 1828) (Schütt, 1970; Ficcarelli, 1984). Moreover, it has been suggested that the extant cheetah should be grouped together with all the Plio-Pleistocene specimens as representing one species (Turner and Antón, 1997). However, consideration of newly discovered fossils from the North African site of Ahl Al Oughlam (Morocco), dated to about 2.5 Ma (Geraads, 1997), alongside those from the East Asian loess deposits around Longdan (Gansu, North China) of 2.55 to 2.16 Ma (Qiu et al., 2004), may support the hypothesis that more than one cheetah taxon roamed the vast territories from North Africa to the Eurasian Far East during the Early Pleistocene. A decision based on reliable biological data, as to whether correspondingly distinguishable forms - Acinonyx aicha Geraads, 1997 (from Ahl Al Oughlam) and Acinonyx linxiaensis (Qiu, Deng et Wang, 2004), in addition to Acinonyx kurteni Christiansen et Mazák, 2009 from Longdan - merit the species rank, or whether a subspecies rank might be more appropriate, should be postponed until the fossil record of cheetah evolution is more complete. In the meantime, it seems reasonable to unite the fossil cheetahs under the mutual name A. pardinensis in the widest sense (sensu lato), following a macrospecies concept established by Hemmer et al. (2008). The latter proposal omits extant A. jubatus, in light of its uncertain phylogenetic relationship to the described fossil forms, resulting from inadequate knowledge of Pleistocene sub-Saharan African cheetah evolution. DNA assessment of the recent subspecies has not helped to elucidate the early phylogeographic roots of A. jubatus, pointing to a population split of the extant Asian and African cheetahs no earlier than the Late Pleistocene (Charruau et al., 2011). The elucidation of the history of European cheetahs has been hindered by the long-lasting confusion surrounding the type locality of the taxon A. pardinensis (Croizet et Jobert, 1828). Over the last 50 years, the type mandibular fragment has been attributed to the site of Étouaires (Schaub, 1949; Viret, 1954; Ficcarelli, 1984; Hemmer, 2001), alongside that of another important cheetah fossil, the type specimen of A. pardinensis arvernensis (Croizet et Jobert, 1828). This latter mandible differs so much from the pardinensis morphotype that in the first modern revision Schaub (1949) accepted their assignment as species of two different genera. Later, this A. pardinensis arvernensis jaw was considered to represent a very robust aged male cheetah individual, with a remarkably long molar (Viret, 1954 and later authors). However, the original authors explicitly stated that the A. pardinensis pardinensis type specimen was found at Ardé, about 1 km away from the Étouaires site (Croizet and Jobert, 1828, p. 208). Postcranial cheetah remains from the latter locality were originally misinterpreted as belonging to the genus Megantereon (revised by Schaub, 1939). In fact, they comprise the only other associated foreleg elements, besides the Dmanisi fossils described here. According to the provenance, the Étouaires postcranial elements are part of the A. pardinensis arvernensis series, but are not that of A. pardinensis pardinensis. Chronostratigraphically, the Ardé sediments postdate the Étouaires (MN 16) type layer (Pastre, 2004). A. pardinensis pardinensis, best known by a cranial series from Saint Vallier of c. 2.0 Ma (MN 17; Guérin et al., 2004), was, therefore, a chronological successor of A. pardinensis arvernensis in Europe. The archaic level of cheetah evolution, as represented by the latter, may have persisted in eastern China until 2.0 Ma, as indicated by recent discoveries from the Tuozidong Cave near Nanjing (Jiangsu; Nanjing Museum and Institute of Archaeology, 2007). Compared with the late Lower Villafranchian A. pardinensis arvernensis foreleg from Étouaires, the Dmanisi foreleg is differentiated by its noticeably more robust humerus and radius structure. The former appears to be more advanced, i.e. A. jubatus-like. In fact, this morphological shift may not have any taxonomic significance, but could be a result of sexual dimorphism, the Dmanisi cat being a very strong male and the Étouaires one a slender female. Such a sexual assignment is strongly supported by body mass estimations of only 70 kg for the Étouaires humerus, rather than indicating differences of taxonomic relevance. Body mass estimations based on these metacarpals, which are thought to belong to the same small individual (Argant, 2004), result in 80 kg for the MC III and 60 kg for the MC V. As the MC V leads to an underestimation of body mass (see Section 4.3) the latter value fits with a possible female cheetah individual of around 80 kg. A distal humerus fragment from the early Middle Villafranchian (c. 2.4 Ma) locality of Liventsovka (Khapry Faunal Complex) near Rostov-onDon (southern Russia) suggests a female of around 60 kg. In contrast, an upper carnassial from the similarly dated site of Morskaya, obviously belonged to a very large male individual (Sotnikova et al., 2002). These Khapry cheetahs may be assigned to A. pardinensis pardinensis, as should the Middle Villafranchian cheetah from Kuruksaj in Tajikistan, which was originally named Acinonyx pamiroalayensis Sharapov, 1986 (Hemmer, 2001; Hemmer et al., 2008). A 100 kg male as estimated for the Dmanisi cheetah, would be in accordance with A. pardinensis pardinensis male body masses of around 100 kg and corresponding female ones of 60e80 kg, but does not argue for its taxonomic affiliation. Corresponding foreleg elements from Early Pleistocene northern Chinese cheetahs A. pardinensis linxiaensis (Qiu et al., 2004), its chronological successor A. pardinensis pleistocaenicus, and A. pardinensis kurteni (Christiansen and Mazák, 2009) have not yet been recorded. A. pardinensis pleistocaenicus appeared in the western Eurasian 1.05 Ma old Epivillafranchian locality of Untermabfeld in Germany (Hemmer, 2001; Kahlke, 2006), only slightly later than its youngest record in the east, at around 1.25 Ma (Qiu, 2006). Two metacarpals from that site, MC III and V (Hemmer, 2001), are much larger than those of the Dmanisi specimen and indicate a huge, surely male, cat. The MC III based body mass estimate is around 140 kg, matching an identical result based on a femur probably from the same individual (Hemmer, 2001). The MC V estimation of around 110 kg may be w20% too low (see Section 4.3). The lack of cranial cheetah remains from Georgia, along with the lack of sufficient comparable fore limb elements from other localities, prevent a more precise diagnostic affiliation of the Dmanisi cheetah within the genus Acinonyx than to that of A. pardinensis, the macrospecies proposed for all Plio- and Lower Pleistocene cheetahs. 5.2. The role of the cheetah in the Dmanisi palaeo-ecosystem The discovery of such a powerful cheetah has helped to close an important ecological gap in the Dmanisi palaeo-ecosystem. This gap had resulted from the complete lack of evidence for a specialized hunter and carcass producer in the open palaeo-landscape around the site, where the existence of hyena (carcass destroyers) had already been established (see Section 3.2). As A. pardinensis (s.l.) shows the exact same structural specialization as the extant A. jubatus, comparable functional use is assumed. This includes hunting as a sprinter in open landscapes, as well as suffocation killing by sustained jaw clench. Preference for this killing method is supported by a tooth-row characterised by rather short and blunt canines followed immediately by high-crowned premolars, which would have provided a steady row of sharp grasping points (Hemmer, 2001, 2004, 2007a). Prey focus mass (PFM, kg) is defined as the modal class that constitutes the prey size preferred by a particular cat species, and is highly dependant on the body mass (BM, kg) of a felid species (method as described by Hemmer, 2004, 2007a; predictor equation: log PFM ¼ 1.917 BM e 1.71; r ¼ 0.96, p < 0.001). Testing the PFM method for the actual African cheetah reveals the peculiar situation of the highly specialized cat. Its mean body mass of 43.2 kg (Marker and Dickman, 2003, Table 2, 3) corresponds to a calculated prey mass value of 26.6 kg, i.e. the PFM class of 20 kg. The mean body mass of the African cheetahs’ preferred prey species, if calculated with three-quarters of mean female body mass in order to take account of calves and sub-adults eaten, was found to be 27.3 4.8 kg (Hayward et al., 2006). This agrees well with the PFM predicted by the body mass. Analysis of the actual prey body mass shows that Namibian male cheetahs prefer to prey on the calves of large antelopes (c. 16 kg), while the females select the smaller antelopes (near 10 kg) (Marker et al., 2003). These figures are only around half of what has been estimated by a prey species age mix procedure and their mean fits the next lower size class (10 kg) on the PFM scale. This agrees with the observation that in addition to body mass, the size of canines also fundamentally influences prey size preference (Hemmer, 2004). Based on the upper canine length predictor equation, the PFM of Namibian cheetahs (Marker et al., 2003) can indeed be calculated to the 10 kg class. In view of these results, the PFM method has been used for the Dmanisi cheetah. A body mass of 100 kg, as estimated for this specimen, in turn suggests that this cat would have focused on prey within the 100 kg class. Taking into account its typical cheetah specialization, this estimated prey body mass value probably indicates the upper limit for the Dmanisi cheetah, rather than its main focus. The 50 kg class of prey mass probably describe its prey target group more realistically. Animals of appropriate body mass are well represented within the Dmanisi large mammalian assemblage, as calves and foals of larger sized species also need to be considered. Under these prerequisites, a considerably higher rate of hunting success compared to other species of the cat guild must be assumed for the Dmanisi cheetah, in parallel with the extant animal (for A. jubatus see Schaller, 1972; Kingdon, 1977; Nowell and Jackson, 1996). This would have qualified the Dmanisi cat to take its place as the top carcass producer within the carnivore community. Occasional loss of prey through encounters with predators ranking higher in the interspecific hierarchy, such as Homotherium crenatidens, Megantereon cultridens and Panthera onca (Hemmer, 2000, 2001, 2004), was presumably tolerable for this highly specialized species. For the extant A. jubatus, Schaller (1972) analyzed 238 kills in the East African Serengeti and found a loss of 12%: twenty kills were taken by lions, eleven by hyenas, and one by a leopard before the cheetah was finished with the meal. The important role of kleptoparasitism has been widely discussed for the extant cheetah (Durant, 2000; Mills et al., 2004; Hayward et al., 2006). A female with cubs seven-eighths fully grown can even be dispersed from a carcass by a subadult spotted hyaena (Crocuta crocuta) (Caro, 1982), indicating the low rank of the cheetah within the extant carnivore guild, independent of the social behaviour of the dominant species. In the Dmanisi ecosystem, the cheetah would, therefore, also have been a principal target species for meat robbery. The carnassial morphology of A. pardinensis in general points to its specialization on fresh meat, just like A. jubatus. Substantial parts of larger kills are left uneaten by cheetahs. The hunting activity of the 100 kg Dmanisi cheetah, focusing on 50e100 kg prey animals, whilst needing an average daily food intake of about 4.5 kg (for food intake prediction from body mass see Hemmer, 2004, 2007a), will have led to the involuntary supplying of other carnivorous competitors with fresh kills, or with carcasses in different stages of consumption. In addition, the Dmanisi cheetah will also have provided significant leftovers for carcass destroyers with bone-crushing capabilities. The following model calculation helps to illustrate the situation. Extant solitary cheetahs make around 150 kills a year (compilation by Schaller,1972). Using a minimal prey focus mass of 50 kg as the mean mass of an A. pardinensis kill, and assuming a similar kill frequency for the latter as estimated for A. jubatus, a total of around 7500 kg of prey a year, seems a realistic hunting estimation for the Dmanisi cat. Cheetahs centre on fresh meat, heart and liver (Schaller, 1972). Taking the example of extant fallow deer (Dama dama), together, these parts of a carcass represent around 40% of live mass (calculated at the base of data compilation by Reinken, 1987). In a scenario involving a 100 kg solitary Dmanisi cheetah individual, around 3000 kg fresh meat of prey would have been produced in a year. Using an estimation of 4.5 kg for average daily food intake, the cheetah itself may have consumed at least 1500 kg, though less than 2000 kg, of this annual meat haul. Consequently, based on the cheetah’s hunting activity, more than one ton of pure meat each year would have been available to other consumers, in addition to around 4.5 tons of leftovers, including bones, hide and intestines. When considering the Dmanisi ecosystem as a whole, the cheetah must have been significantly more suited to the role of a potential prime fresh meat supplier than the dirktooth, the sabretooth or the jaguar, as it would have been a potential host for kleptoparasitism by other carnivorous mammals, which could dominate it in the interspecific hierarchy. By producing considerable amounts of fresh meat and other nutritious leftovers, the hunting success of Acinonyx would have benefited other meat consumers. Even for sympatric Homo, it might have been an occasional factor in their subsistence strategy. The available archaeological evidence indicates that Dmanisi Homo had to some extent early access to carcasses following a kill, and filleted the meat before any large carnivore was able to leave traces of consuming activities (Lordkipanidze et al., 2007, supplementary information; Tappen et al., 2007). This evidence fully agrees with the scenario of Dmanisi man as a potential hunter, as well as a potential confrontational scavenger, who was willing to drive off large felids from their fresh kills (Hemmer, 2000). 6. Conclusions The detection of a large and stoutly built cheetah adds a new element to the extensive list of the Dmanisi palaeofauna. This felid would have successfully filled the ecological niche, previously thought to have been unoccupied, for a specialized hunter and carcass producer in the dry, open areas of the Dmanisi palaeo-landscape. The position and taxonomic affiliation of this large cat within the evolutionary history of the macrospecies A. pardinensis s.l. could not be ascertained in this study due to the lack of cranial elements. However, a shared range with A. pardinensis pardinensis might be possible. The existence of an approximately 100 kg cheetah at Dmanisi, with a hunting specialisation just like that of the much smaller extant A. jubatus, supports the idea of this species as having been a potential prime fresh meat supplier in Palaearctic Early Pleistocene ecosystems, in preference to any other felid.
The original document of Croizet et Jobert, which is the first literature describing giant cheetah; it is in french.
Research on the fossil bones of the department of Puy-de-Dome The above sentence is hyper linked^
Reconstruction of the Giant cheetah by Dr.Hemmer and Dr.Kahlke
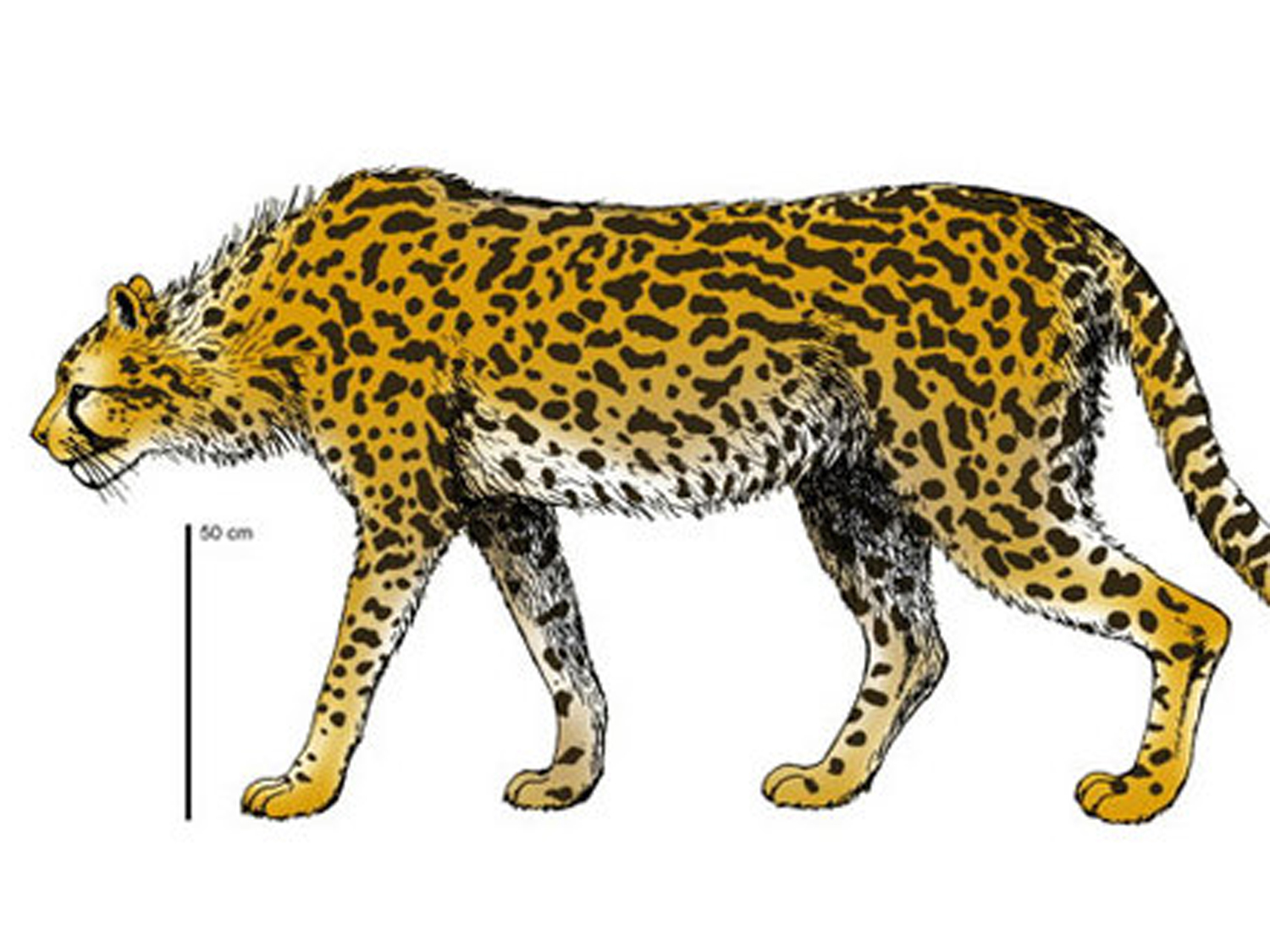 *This image is copyright of its original author
Life in the Slow(er) Lane: Revisiting the Long-Lost Giant Cheetah
For the most part, a cat is a cat is a cat. Large or small, domestic or wild, most are agile ambush predators that subsist almost entirely on meat. But then there’s the cheetah. It’s a cat that hunts like a dog. The felid’s claws are more like cleats than the retractable armaments of its relatives, its nasal cavity is enlarged to house the soft tissues necessary to keep it cool while sprinting, and, as the fastest land mammal, the cat relies on speed to chase down and trip up fleeing antelopes and gazelles. Cheetahs are so different, in fact, that figuring out when they adopted this speedy lifestyle has been clouded by the imagery invoked when we apply the term “cheetah” to their fossil relatives. Take Miracinonyx, for example. The two species of this svelte fossil cat have often been called “American cheetahs” on the basis of their bones, and even inspired the idea that cheetahs first evolved in the New World rather than the Old. But this idea fell apart. Skeletal and genetic clues have shown that Miracinonyx was more closely related to cougars than cheetahs, and there’s some doubt about whether this cat rocketed over open ground after pronghorn or stalked steep rock walls and caves. The title of “American cheetah” invokes scenes of speed, but, even if the carnivore did so, this cat was a false cheetah. The modern cheetah’s true fossil relatives started off in Africa about three million years ago and eventually dispersed through Eurasia before leaving only one species behind. But even though they were more closely related to today’s Acinonyx jubatus than other cats, the prehistoric forms didn’t live and hunt just like the living one. In fact, as Sorbonne University paleontologist Denis Geraads recently concluded, the cheetah as we know it today is a relatively recent evolutionary spinoff. The focus of Geraads’ study was a skull of Acinonyx pardinensis found in France. This species ranged from Spain to Georgia about 2.4 million years ago, and previous paleontologists had concluded that the felid’s skull was already quite similar to that of the modern cheetah. Through using a technique called geometric morphometrics to compare the cat’s skull shape to that of other felids, however, Geraads found that Acinonyx pardinensis was not simply a big version of its living relative. The cat’s skull shape more closely resembled that of a cougar than the more specialized short, deep form of today’s cheetah. In short, the giant cheetah had a skull more like that of other pantherine cats, and the distinctive profile of the modern species evolved much more recently. Two skulls and a jaw found in Italy bolster Geraads’ argument. Uncovered at a site in Pantalla, Italy and described by Perugia University paleontologist Marco Cherin and coauthors, these Aciononyx pardinensis skulls had some cheetah-like traits – such as a shorter relative length and enlarged nasal openings – but they also retained some traits of their ancestors, such as a high keel on the back of the skull for greater jaw muscle attachments. Acinonyx pardinensis was its own cat, with a skull intermediate in shape between today’s cheetah and its more cougar-like ancestors. So what does this mean for how the extinct cheetah hunted? On the basis of muscle reconstructions threaded on CT scans of the Pantalla skulls, Cherin and colleagues tentatively suggest that the extinct cheetah had enough biting power to crush neck and skull bones like jaguars and cougars do. (Modern cheetahs, by contrast, often kill prey with a suffocating death grip on the throat.) More than that, Cherin and coauthors point out that Acinonyx pardinensis weighed about 176 pounds – twice as heavy as modern cheetahs – and that the relatively slender proportions of the cat’s postcrania cannot be immediately taken as evidence that it was a fast runner. The skeletons of snow leopards show some striking similarities to those of cheetahs, the researchers note, even though the two cats occupy very different habitats and hunt in very different ways. Despite its relationship to the modern cheetah, Acinonyx pardinensis was more like a typical big cat. The term “cheetah” isn’t inaccurate for Acinonyx pardinensis. The fossil cat surely belonged to that lineage. But the title has also obscured how different this felid truly was. The exceptional nature of the modern cheetah has masked the unique nature of its fossil relatives. If we can remove that bias and understand fossil species in the context of their own time, we gain more than a richer understanding of the past. We earn a deeper appreciation for the vast changes that made our modern megafauna what they are. References: Cherin, M., Iurino, D., Sardella, R., Rook, L. 2014. Acinonyx pardinensis (Carnivora, Felidae) from the Early Pleistocene of Pantalla (Italy): predatory behavior and ecological role of the giant Plio-Pleistocene cheetahAcinonyx pardinensis (Carnivora, Felidae) from the Early Pleistocene of Pantalla (Italy): predatory behavior and ecological role of the giant Plio-Pleistocene cheetah. Quaternary Science Reviews. doi: 10.1016/j.quascirev.2014.01.004 Geraads, D. 2014. How old is the cheetah skull shape? The case of Acinonyx pardinensis (Mammalia, Felidae)How old is the cheetah skull shape? The case of Acinonyx pardinensis (Mammalia, Felidae)How old is the cheetah skull shape? The case of Acinonyx pardinensis (Mammalia, Felidae). Geobios. doi: 10.1016/j.geobios.2013.12.003
Important remarks on the evolution of the giant cheetah from Cherin et al 2014:
"Evolutionary relationships among all these forms, as well as between Old and New World forms, are still cloudy. At least two taxa were suggested to be the possible ancestor of cheetahs: the puma-like cat Puma pardoides from Eurasia (Kurtén, 1976) and Panthera crassidens from Africa (Petter and Howell, 1976). The latter however should not be considered because the holotype was demonstrated to be a composite specimen (Turner, 1990). The American cheetah M. inexpectatus probably reached North America in the Late Pliocene and could be ancestral to both M. trumani and the extant cougar Puma concolor (Van Valkenburgh et al., 1990). Miracinonyx inexpectatus and A. pardinensis seem to appear approximately at the same age. Although the origin of A. pardinensis is still unclear, Hemmer et al. (2011) supposed that European and Asian forms followed a separate evolutionary history during the Early Pleistocene: while in the West A. p. pardinensis was probably a chronological successor of A. p. arvernensis during the early to middle Villafranchian transition, in the East the primitive A. p. linxiaensis was substituted by A. p. pleistocaenicus, which invaded Europe in conjunction with the climatic aridification at the beginning of the latest Villafranchian (Spassov, 2011)." I will post most more weight data on pardinensis than I have before very soon.
Data on Giant Cheetah subspecies:
 *This image is copyright of its original author
Body mass data on the giant cheetah:
Body mass Georgia A male giant cheetah in Dmanisi was estimated to weigh around 100 kg. (3) France A Female giant cheetah from Étouaires had an estimated body mass of 70 kg, possibly a different female specimen from the same place had an estimated body mass of 80 kg. A male cheetah from Étouaires had an estimated body mass of 120 kg. A female giant cheetah from Cote d’Arde had an estimated weight of 70 kg; a female from Saint Vallier was estimated to weigh 60 kg. (1,2) Germany Two male giant cheetahs from Untermassfeld had an estimated mean body mass of 125 kg (n=2), they were estimated to weigh 110 kg and 140 kg respectively; the latter is the largest giant cheetah specimen found as of yet. A female cheetah from Mosbach was estimated to weigh 60 kg. (1,2) Italy A male giant cheetah from Casa Frata had an estimated body mass of 110 kg. A female from Pantalla was estimated to weigh 80 kg; a female from Montopoli was estimated to weigh 60 kg; a female from Villafrana d’Asti was estimated to weigh 70 kg. (1,2) India A male giant cheetah from the Siwalik Hills was estimated to weigh 110 kg. (2) China A male cheetah from Longdan was estimated to weigh 90 kg. (2) Morocco A male giant cheetah from Ahl Al Oughlam was estimated to weigh 100 kg. (2) Spain A female in Fonelas had an estimated body mass of 60 kg. A male from Villaroya had an estimated body mass of 90 kg. (2) Total average: Males: 110 kg (n=8) Females: 70 kg (n=8) References: 1. Evolution of community composition in several carnivore palaeoguilds from the European Pleistocene: the role of interspecific competition. 2. Acinonyx pardinensis (Carnivora, Felidae) from the Early Pleistocene of Pantalla (Italy): predatory behavior and ecological role of the giant PlioePleistocene cheetah 3.The cheetah Acinonyx pardinensis (Croizet et Jobert, 1828) s.l. at the hominin site of Dmanisi (Georgia) - A potential prime meat supplier in Early Pleistocene ecosystems 4.The Carnivores of the late Pliocene site of Saint-Vallier (Dr6me, France) 5. Argant, A., 2004. Les Carnivores du gisement Pliocène final de Saint-Vallier 6. Evolution of the extinct Sabretooths and the American cheetah-like cat 7. A record of cheetah scavenging in the Serengeti 8. Phylogeography, genetic structure and population divergence time of cheetahs in Africa and Asia: evidence for long-term geographic isolates. 9. Recherches sur les ossemens fossiles du département du Puy-de-Dôme 10. Les faunes de grands Mammifères de la Grotte du Vallonnet Roquebune-Cap-Martin, Alpes-Maritimes 11. Nuove ricerche sui Felini del Pliocene italiano 12. The Villafranchian cheetahs from Tuscany and remarks on the dispersal and evolution of the genus Acinonyx 13. The paleobiology of the extinct venomous shrew Beremendia (Soricidae, Insectivora, Mammalia) in relation to the geology and paleoenvironment of Dmanisi (Early Pleistocene, Georgia). 14. Archidiskodon gromovi Garutt et Alexeeva - der älteste Elefant der Mammutlinie in Eurasien 15. Carnivores du Pliocène terminal de Ahl Al Oughlam (Casablanca, Maroc) 16. Gallogoral (nov. gen.) meneghinii (Rütimeyer, 1878), un rupicapriné du Villafranchien d’Europe occidentale 17. Le gisement pliocène supérieur de Saint-Vallier (Drôme, France): synthèse biostratigraphique et paléoécologique
Visual comparisons concerning the giant cheetah (made by me)
Figure 1: Visual Comparison between Giant Cheetah and Cheetah (average males) *This image is copyright of its original author Figure 2: Visual Comparison between Human and Giant Cheetah (Average males) *This image is copyright of its original author Figure 3: Visual Comparison between Giant cheetah and Asiatic cheetah humerus *This image is copyright of its original author Figure 4: Visual Comparison between Giant cheetah and Asiatic cheetah humerus *This image is copyright of its original author
Acinonyx pardinensis (Croizet et Jobert) remains from the Middle Villafranchian locality of Varshets (Bulgaria) and the Plio-Pleistocene history of the cheetahs in Eurasia
ABSTRACT The article decribes and discusses remains of late Pliocene (middle Villafranchian) cheetah, Acinonyx pardinensis (Croizet et Jobert) from Varshets, N.-W. Bulgaria. It is accepted that three chronoforms A. pardinensis pardinensis, A. p. pleistocaenicus and A. p. intermedius succeed in Europe from the Early Villafranchian (Etouaires, approx. 2.6 Ma) till the Middle Pleistocene. Remains are scarce and at this level of knowledge we could accept the recently proposed subspecific status of all these forms but their taxonomical relations could be rather more complicate. The remains from Varshets correspond to the earliest form A. pardinensis pardinensis which inhabited Eurasia during the late Early and the Middle Villafranchian and the beginning of the Pleistocene. Other European remains of A. p. pardinensis are known only from Spain, France, Italy and the Azov region of Southern Russia. It seems that then the species occupied only the southern part of the European continent (the Mediterranean-Balkan-Northern peri-Pontic area), and from there it probably spread till Central Asia (Tajikistan). Such a distribution supports the concept of the faunal entity of South Europe in Villafranchian time as well as the theory for the Central Asian influence of this fauna mainly trough the Balkans by two ways: via Bosphorus and via the peri-Pontic area. According to the paleontological data the fossil Acinonyx does not reach Central Europe (Austria and Germany) before the Epivillafranchian, possibly after new waves of dispersal of another, Early Pleistocene form of Central Asia. The relationship of the Middle Villafranchian A. pardinensis with the rather contemporaneous forms from China (Hezheng) “Sivapanthera” linxiaensis and A. kurteni and from Siwaliks is not clear, but the Chinese forms must represent taxa different from A. pardinensis. Acinonyx s. str. is characterized by strongly domed and enlarged frontal area, shortened skull and downwards inclined neurocranium. The new data argue the supposition of its Eurasian origin at the end of the Ruscinian/ beginning of the Villafranchian. Introduction The well known Middle Villafranchian locality of Varshets, N-W Bulgaria, has yielded more than 150 vertebrate species: about 20 amphibian and reptile species (M. Böhme, N. Tzankov pers. comm.), 74 bird species (BOEV, 1999, 2000), 37 species Micromammalia (POPOV, 2001) and 21 species Macromammalia (Spassov, 2000 and additional pers. data). This is evidently the richest Pliocene locality of vertebrate fauna in Europe. The mammal megafauna of this locality demonstrates an apparent similarity with localities typical for the late Pliocene – Middle Villafranchian, such as SaintVallier and Chillhac (France), La Puebla de Valverde (Spain), etc. Varshets is placed, after its macromammal fauna, in the first half of the NMQ17, between Roccaneyra (possibly close to) and St. Vallier. The appearance of the fauna of Varshets shows great similarity to that of the European-Mediterranean localities but unlike most of the well known Mediterranean faunas, that of Varshets also shows a connection with the faunal associations of Middle- and E-Europe. The relationship with the Middle East is evidently strong, too. A tendency of prevalence of taxa with S-European and Middle East distribution emerges. This, in fact, appears to be quite logical bearing in mind the geographical position of the locality and the clear indications of an influence of Asian elements (SPASSOV, 2000; 2003). The distribution of the macromammalian species from Varshets in different habitats shows that the physiognomy of the landscape is determined chiefly by forest-steppe/mosaic biotopes or open forests. The open forests (park-like forests) and shrubs prevail. The analysis of the micromammalian species leads to similar conclusions for a temperate, mild climate and a mosaic landscape with forests and meadows prevailing over typical steppe vegetation (POPOV, 2001). The data from the analysis of the ornithofauna are similar. The birds of the open spaces are 63 % of the number of the forest birds, but dominant are the birds of the mixed (mosaic) environments, especially the phasianids (BOEV, 1999). Till now Acinonyx pardinensis Croizet et Jobert is only mentioned in the check-list of Varshets (Spassov, 1997; 2000; 2003). The fossil remains of cheetah in Europe are still rare, that determines the importance of the description of the Acinonyx remains from Varshets. Material (Coll. Of the NMNHS): Upper canines (FM 849; FM 851), upper carnassials – P4 (FM 953; 954; 955, all are possibly from three female individuals), astragalus FM 873; calcaneus FM 878 (the astragalus and calcaneus are most probably from one and the same ind.); I phalanx FM876 (Fig.1). We could refer to this species also the following remains: metapodials (FM 870; 879); II phalanxes (FM 874; 875; 877); III phalanx (FM 983) and two bones of a juvenile individual: a symphysis fragment (FM 862) and a calcaneus (FM868). The remains from Varshets, belonging to at least 4 individuals, make the cheetah the most numerous carnivore species in the taphocoenosis of the locality. Description and comparison The canines (FM 849; 851) have the characteristic cheetahlike crown shape: relatively short and thick. The typical felid longitudinal furrows are visible on the labial surface. Normally these furrows are absent on the A. jubatus canines. The aforementioned furrows are also not visible on the specimens from the Epivillafranchian cheetah from Untermassfeld (Hemmer, 2001), but exist, weakly marked on the canines of the specimens from St. Vallier, as well as in the lower canines of A. pardinensis from the MNQ18 of Fonelas (Spain) (Garrido, 2008). Unlike most felids however, those furrows on the Varshets canines are relatively weak and short. Probably this felid character disappears secondary in cheetah and could be more frequently manifested in the larger teeth of the older populations of the genus. The canines correspond in their size to the compared specimens of A. pardinensis from St. Vallier (MNQ17) (table1). Upper P4s are characterized by a strong reduction of the protocone which shapes no point and is very slightly protruding in lingual direction. It looks more like a prolongation of the lingual root of the tooth at the crown level. The root itself is closely pressed to the main body of the tooth. Such morphology is typical for cheetahs (Teilhard De Chardin & Leroy, 1945; Viret, 1954; Sotnikova, 1989). In this respect, the P4 differs clearly from the P4 of the recently described species from the Late Pliocene of China, named Acinonyx kurteni (see below). The size of the varshets P4 is statistically larger than the Recent A. jubatus. The preparastyle of the Varshets specimens is only vestigial, clearly less developed than in the recent form, which explains why the mesial paracone border is more oblique than in the recent species. The protocone of the Recent species is slightly more distally displaced. In the position of the protocone (displaced mesially), the Varshets specimens also correspond to another specialized form that is close to Acinonyx and mainly to Puma – the North American Miracinonyx inexpectatus (= M. studeri) (JWT 563-420664 - cast, col. SI-NMNH). The size and morphology of the Varshets P4s completely correspond to A. pardinensis from the Middle Villafranchian of Saint-Vallier (France). Both the MNQ17 samples, the sample from St.-Vallier and the one from Varshets do not reach the dimensions of the Untermassfeld (Germany) A. p. pleistocaenicus teeth (see Hemmer, 2001). The maximum dimensions of the Late Pliocene N. African A. aicha (Geraads, 1997) are also larger. At the same time, the European cheetahs from St. Vallier and Varshets have slightly thicker upper P4s than the mentioned primitive African species (Table 2). The calcaneus (FM 878) is typically felid (see Gromova, 1960), its size is rather larger than a jaguar and much smaller than a male lion. The sustentacular (inner astragalus) facet is obviously rounded, rounder than in a jaguar or leopard for example. Its articular surface is in a single plane. Plantary from the sustentaculum there is a very strong furrow which usually is not that deep in the noted species of genus Panthera. In the proximal end, on the lateral surface, there is a pronounced elongated fossa. In proximal direction it reaches the proximal edge of the bone (i. e. the edge of the cuboid facet). This fossa is practically missing in lions and jaguars. The bone is larger than the calcaneus of Megantereon and lacks its strong convexity on the lateral surface of the proximal end. Although of approximately the same total length, the Varshets calcaneus differs strongly from Homotherium: it is much more slender, and its distal portion is quite elongated; it lacks also the navicular facet typical of Homotherium (Ballesio, 1963). Significantly larger than the calcaneus of a Recent cheetah (60-78 mm after Gromova, 1960), the Varshets specimen fully corresponds in shape and size to the calcanei of A. pardinensis from Saint-Vallier stored in the MHNL (Table 3) but does not reach the values of the specimen from Untermassfeld. The astragalus is typically felid. Compared to Homotherium (with length 48.3 mm and width 45.5 mm – coll. MNHNP, Lab. de Paléontologie) it is insignificantly longer but much narrower, with a narrower trochlea and differently situated caput. The caput of the astragalus looks “twisted” in ventral view, if compared to other large felids like leopard and jaguar: its maximal diameter is obliquely placed, compared to the noted species of genus Panthera. There is a strongly expressed droplike fossa on the lateral surface of the caputs’ column, with its broader part ending ventrally on the edge of the articular surface of the caput. The elongated sharp end of the fossa touches the small facet for the calcaneus. A similar fossa is present on the astragalus referred by Kurten & Crusafont (1977) (see there the figured astragalus) to A. pardinensis from Puebla de Valverde. In all noted characters and in size (table 4.) the Varshets astragalus corresponds to the astragalus of A. pardinensis from St.-Vallier (Coll. MHNL) but it is smaller than the specimen from Untermassfeld. The Varshets cheetah has morphological and morphometrical differences with the modern species. The postcranial remains are not only of larger size than the recent species, but also have more massive proportions. The fossil cheetah from Varshets was much larger than the Recent species and possibly not so fast. After the visual comparison of the astragalus and the calcaneus with these of different recent large cats with known weight, the body weight could be about 100-110 kg. The average weight of the recent male A. jubatus is somewhat more than 40 kg (Krausmann & Morales, 2005) and could reach no more than 65-70 kg. The upper P4 of the cheetah from Varshets has more oblique and elongated parastyle (as in “Miracinonyx” inexpectatus) than in the recent species. This puts their protocone in an even more anterior position than in recent cheetah – almost at one level with the parastyle. Those differences, despite the individual variations, show an obvious regularity. Having in mind the strong difference in body size (see below), the only statistically larger P4 of the Varshets form represents a rather plesiomorphic stage of the carnassial teeth in relation to the Recent species. Both morphological and morphometrical data show that the discused fossils belong to large cheetahs, fully similar according the existent data to Acinonyx pardinensis (Croizet et Jobert) from the Early Villafranchian to the beginning of the Late Villafranchian of Europe. The morphological and metrical comparison of the fossil cheetahs is difficult because of the scarce material and the body weight comparison is only approximate. If the body mass of the Varshets cheetah (~ 100-110 kg after the postcranial remains) is comparable with the estimated weight of the Villafranchian A. pardinensis (probably males and females) from Les Etouaires (80- 120 kg), St. Vallier (60-80 kg, calculated after tooth length, but A. pardinensis has smaller carnassials related to A. jubatus: see above), Olivola and Casa Frata (90-120 kg) and Dmanisi (around 100 kg), it would be smaller than the Epivillafranchian cheetah from Untermassfeld (110-140 kg) and larger than the Middle Pleistocene cheetah from Hundsheim and Mosbach (60- 90 kg.) (see Hemmer, 2001; Hemmer et al., 2008; Hemmer et al., 2011). Discussion. What is a cheetah? Problems of the taxonomy, the origin and the phylogeography of the cheetahs The time and place of the origin of the genus Acinonyx are still debatable. Africa has been pointed out as one possibility (Sotnikova, 1989). Indeed, there are African finds referred to cheetahs, having a supposed absolute age ca. 4 Ma (Omo Mursi) and more than 3.5 Ma (Laetoli). Those remains, however, are very fragmentary, and their generic belonging is not unequivocal (Turner, 1990). Turner (1990) supposed that cheetahs are immigrants into Africa. Recently, a new cheetah species with some plesiomorphic features compared to A. Pardinensis, such as a less convex and less specialized skull, was found in Africa (Geraads, 1997). The age of this form, Acinonyx aicha Geraads is about 2.5 Ma. In Africa, the oldest unequivocal fossil remains of genus Acinonyx s. str. are as old as 3 Ma (early MN16) and have been found both in Africa (Makapansgat 3) (Turner, 1990). In Europe the oldest A. pardinensis remains are from Perrier-Etouaire, from where comes the type specimen (a mandible: see Schaub, 1949) of the species A. pardinensis Croizet et Jobert. The age of Les Etouaires is not very clear but it is referred to MNQ16b (Steininger et al., 1990) and could be at about 2.5 Ma (Guérrin, 2007) or most probably slightly older, close to the age of Montopoli (Aguirre et al., 1997). The oldest known Asian remains (India – Pinjor, China) could be slightly younger. Recently a new species of cheetah with an estimated age of 2.2-2.5 Ma, that demonstrates a typical cheetah skull shape, but a rather plesiomorphic stage of the upper carnassials, was described in Hezheng area, China under the name of A. kurteni (Christiansen & Mazák, 2009). A. kurteni seems to be an Acinonyx s. str., characterized by advanced skull shape but plesiomorphic tooth morphology. Having in mind the features of this middle Villafranchian Asian specie and the co-existence of another species in Europe, we could suppose the presence of two parallel lineages of cheetahs in Eurasia from the Early Villaftranchian: A. kurteni and A. pardinensis1, and to imagine an Eurasian origin of the genus in Late Ruscinian/Early Villafranchian time. It is accepted that Acinonyx (and possibly Uncia) is a close relative of Puma (Salles, 1992; Van Valkenburgh et al. 1990) (The close affinities of the snow leopard with the Acinonyx-Puma group is contested by recent DNA investigations (Werdelin et al., 2010)). The question of the place and time of the Puma origin is also still unclear. A form morphologically similar to cheetah, the American “cheetah” – Miracinonyx (Van Valkenburgh et al., 1990), that was referred until recently to Acinonyx s. str., possibly shares a common ancestor with Puma from which both, Puma and Miracinonyx diverged, after some opinions, in N. America at about 3.2 Ma ago (Barnett et al., 2005). As Acinonyx, Puma is also known in the Early Villafranchian of Eurasia with a form, that merits subgeneric separation as P. (Viretailurus), having some skeletal and cranial differences from the recent form (personal observations). The oldest Puma localities known are Shamar, Mongolia and Kvabebi, Georgia but also Perrier-Etouaires, France (Hemmer et al., 2004; Kurten & Crusafont (1977) and Red Crag, Great Britain, all of them with Early Villafranchian age. Hemmer et al. (2004) refer some African remains with an age of about 3.5-3 Ma to Puma and suppose an African origin of the genus. Except from Varshets European fossil cheetah are known from about sixteen early Villafranchian to middle Pleistocene localities of France, Spain, Italy, Austria and Germany (See Garrido, 2008 and Hemmer et al., 2008) as follows: Etouaires, Saint Vallier, Senèze, Le Vallonet, Saint-Estève (France); La Puebla de Valverde, Las Higueruelas, Villarroya, Fonelas (Spain); Montopoli, Villafranca d’Asti, Olivola, Casa Frata (Italy); Untermassfeld and Mosbach (Germany); Hundsheim (Austria). A find from Liventsovka (Priazovian South Russia, NMQ17) noted as Acinonyx sp. (Titov, 2008) must represent the same species. After the recent concept (Hemmer et al., 2008) the European fossil cheetahs belong to a single and polymorphic large species or “macrospecies” - A. pardinensis (s. lato). The status of Felis arvernensis Cr. et Job., described, as well as A. pardinensis (Cr. et Job.) from Etouaires is still debatable. Some authors express the opinion that this cat has specific differences from A. pardinensis (see Heintz et al., 1974; see also Hemmer et al., 2008). It is difficult to decide from one hemimandible only if this is a separate taxon or an aberrant individual of A. pardinensis from Perrier - Etouaires. Possibly, however, Viret (1954) was right to consider this taxon a synonym of A. pardinensis, and this seems a more parsimonious hypothesis. Indeed, the type mandible has some atypical features such as the elongated distal part of P4, the shape of the paraconid (a specific occlusion) etc. At the same time, a number of characters such as the short massive canine, the short diastema, the rather massive symphysis and the position of P3 indicate cheetah affinities (Spassov, 1999). The history of cheetahs in Europe is still unclear, due to the scarce material. I will follow Hemmer (2001) and Hemmer et al., (2008) who proposed a taxonomic concept according to which the fossil cheetahs of Europe could be distributed in three successive taxa with subspecific rank: A. p. pardinensis (Cr. Et Job.), for the Early Villafranchian till the Pleistocene beginning (Etouaires – Casa Frata), A. p. pleistocaenicus (Zdansky) for the Epivillafranchian form (Untermassfeld): a migrant from Asia) and A. p. intermedius (Thenius) for the Middle Pleistocene cheetah (Hundsheim and Mosbach). At the same time the Cheetah evolution could be much more complicated. Having in mind the considerable time span between the first and the last representative of the species on the continent we could suppose specific differences between the separate successive forms. Besides the probable size differences, some morphological differences support this supposition: Hemmer (2001) and Hemmer et al., (2008) note some morphological differences in the mandibular symphysis between A. pardinensis pardinensis (the Etouaires – Olivola form) on one hand and the Epvillafranchian (Untermassfeld) and the Middle Pleistocene (Hundsheim, Mosbach) forms on another hand. If we judge after the recent carnivores, such kind of features that concern not only size and proportions but also the skull morphology could be regarded as criteria for specific distinction. After this second taxonomic possibility A. pleistocaenicus could be regarded as a separate Asian species, that invaded Europe during the strong aridification at the beginning of the Epivillafranchian. The Varshets remains fill the knowledge gap on the dispersal of A. pardinensis (especially of “A. p. pardinensis” sensu Hemmer et al., 2008) in the Villafranchian of Europe and show its presence not only in the Western Mediterranean but also in Southeastern Europe. We could conclude now, after the distribution of the known cheetah localities, that in the time interval between the Early Villafranchian and the beginning of the Pleistocene this cheetah occupied only the southern part of the European continent (the Mediterranean-Balkan area). From there, it was probably spread through the peri-Pontic area (South Russia - Liventsovka: Sotnikova, 1989 and Dmanisi - Georgia: Hemmer et al., 2011) till Central Asia: Tajikistan (see Sotnikova, 1989) and in case that Felis brachygnathus Lydekker, 1884 (= Sivafelis brachygnathus (Lydekker) sensu Pilgrim, 1932) from Pinjor (Siwalik) represents the same species of cheetah, it could be spread also till India. Such a distribution supports the idea of the specific character of the South European fauna in Middle Villafranchian and of the strong Central Asian influence on this fauna trough the Balkans (Spassov, 2000; 2003). According the paleontological data the fossil cheetah does not reach Central Europe (Austria and Germany) before the Epivillafranchian and this is possibly related to a new wave of dispersal from the east, related to the continuation of the aridification. The assignation of the scarce Indian (Pinjor) material to A. pardinensis is debatable. Qiu et al. (2004) are probably right to separate their new species Sivapanthera linxiaensis from Acinonyx. The skulls of this probably Middle Villafranchian taxon that is again (as A. kurteni) from Longdan, Hezheng area (China), figured in Qiu et al. (2004) differ from Acinonyx mostly by the less domed frontals, the large infraorbital foramen, the strong and elongated sagital crest, the more elongated muzle, the pinched caudal portion of the muzzle (the maxillary bones) on both side of the nasals. With all these features this new taxon shows similarities with Puma and could be regarded as a member of the Acinonyx stem-group. Its relations with Miracinonyx could represent certain interest. At the same time, the close affiliation of the later form “Cynailurus” pleistocaenicus Zdansky with Sivapanthera linxiaensis does not seem to be strongly supported by the paleontological facts and I will follow Hemmer et al. (2008) in the interpretation of this taxon as a trough cheetah, that migrated in the Epivillafranchian in Europe. The recent fossil material described in Central Asia complicates the problem of the definition of the Acinonyx diagnosis: several forms of Pantherasized cats with (1) short canines and (2) relatively developed frontal sinuses as well as with different degree of expression of the typical cheetah/cheetahlike facial and carnacial features have existed in a very large territory in the Early/ Middle Villafranchian: 1. forms with relatively flattened frontals; medium elongated/shortened skull; large (?) infraorbital foramen, but P4 protocone, with a cheetah-like trend to reduction (Acinonyx aicha, “Sivapanthera” linxiaensis; Miracinonyx); 2. forms with well to strongly domed and enlarged, cheetah-shaped frontal area and shortened skull; but with 2.1.: unreduced P4 protocone (A. kurteni) or 2.2.: reduced protocone (A. pardinensis). We could mention that the reduction of the P4 protocone is a feature that has appeared more than once in the felid evolution and this trend could appear in parallel especially in closely related forms. Possibly the most peculiar features of the Acinonyx s. str. skull is the visibly bulged frontals, related to the strong development of the frontal sinuses. The development (to a different extent) of the frontal sinuses and the bulging of the frontal area represent synapomorphy of the closely related Puma, Acinionyx and Uncia but the extremal stage this development is reached in Acinonyx. The specific inclined position of the neurocranium of Acinonyx in respect to the palatal plane and the related more oblique position of foramen magnum, as well as the strong vertical elevation of the fronto-orbital region in this felid correspond to the important role of vision and are typical features of the cheetah. These features could be regarded as autapomorphies of Acinonyx as adaptation to a mode of life in open landscapes. Post scriptum from the author After this paper was in print Deng Tao’s publication (2011) came out. This paper revealed the news that the skull (type and single specimen) of this newly described species from China (Christiansen & Mazák, 2009) is a fossil forgery. Deng (2011) especially notes that the zygomatic arch, the occipital part, the sagital crest, the basiooccipital area and parts of the parietal area (above all the posterior part of the skull) bear the traces of artificial modeling. It is possible that the cheetah-like domed shape of the skull roof has not been affected by the human interference (see the answer of Christiansen & Mazák cited in Deng Tao (2011), however we could not be sure in the natural appearance of the skull’s contour of this Chinese specimen (Deng Tao, pers. comm.). Neither the age, nor the location of the remains are clear; it could have been from Longdan (2.2-2.5 Ma), and neither is that sure. For this reason the shape of the skull and the name and age of the location cannot be used as firm arguments concerning some of the theses offered in my publication, and in particular: • the existence of “two parallel lineages of cheetahs in Eurasia from the Early Villagranchian: A. kurteni and A. pardinensis “. For this reason the probability, put forth in the paper that the cheetahs rather originated in Eurasia cannot be launched with priority before the version that they are African in their origins. The issue is still awaiting its solution. The existence in the Middle Villafrachian of forms with strongly domed and enlarged, cheetah shaped frontal area and shorten skull, but with unreduced P4 protocone cannot be accepted for certain. It is quite possible that the strongly domed skull was a relatively late apomorphy with cheetahs, dating as late as the Pleistocene. AKNOWLEDGEMENTS The comparison of the material is carried out in different museums’ collection in the 90s with the support of the Bulgarian Academy of Sciences – Dept. of International Collaboration, the Bulgarian Ministry of Science and Education as well as the Embassy of France in Sofia. The author is very grateful to Zhanxiang Qiu (Inst. of Vertebtrate Paleontology and Paleoanthropology, Beiging) for the helpful discussion, to the late Leonard Ginsburg (MNHN-Paris), Michel Philippe (MHNL) and Steve Jabo (Smithsonian Institution, NMNH, Washington D.C.) for the access to the fossil collections and the useful comparative photos sent. Thanks to Jorge Morales as well as two the anonymous reviewer for the very useful remarks and comments that improved the manuscript. |



| Users browsing this thread: |
| 1 Guest(s) |